РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ - Рео - та аденовірусна інфекції і їх вплив на ефективність вакцинопрофілактики інфекційного бронхіту птиці
Результати серологічного моніторингу птиці промислових птахогосподарств, синантропних і диких птахів щодо адено - і реовірусної інфекцій, ефективність вакцинопрофілактики інфекційного бронхіту курей. Ефективність вакцинопрофілактики вірусних інфекцій в промисловому птахівництві залежить як від правильного підбору програм вакцинації, так і врахування епізоотичної ситуації у регіоні. Для цього необхідним є впровадження моніторингових досліджень птиці щодо визначення спектру латентних вірусів, що циркулюють у господарстві.
Проведений нами серологічний моніторинг синантропної, дикої та свійської птиці виявив наявність у неї антитіл у титрах різного рівня до адено-, рео - та вірусу ІБК. До 80-100% синантропних птахів (горобці, голуби, ворони), що реагували до адено-, рео - та вірусу ІБК, встановлено у Сватівському, Станично-Луганському, Краснодонському та Перевальському районах Луганської області.
Високий титр антитіл (4,0 Log2) до реовірусу та аденовірусу виявлено у свійської птиці (кури, гуси) у Біловодському районі Луганської області, серед синантропних птахів (3,6 Log2 і 3,4 Log2, Відповідно), які мешкають на території промислових птахогосподарств Краснодонського району, ВАТ "Агроукрптаха" та ВАТ "Сімейкінське". Аналогічна ситуація спостерігалася також серед синантропних птахів ВАТ "Червоний прапор" Перевальського району та СТОВ "Авіс" Лутугінського району Луганської області (титр антитіл до рео - та аденовірусів - 3,0 Log2).
Серед диких птахів (граки, качки) антитіла до зазначених вірусів на рівні 3 Log2 реєстрували у Станично-Луганському, Марківському та Попаснянському районах Луганської області.
Найбільш високий рівень антитіл до вірусу ІБК встановлено у Станично-Луганському, Кременському та Попаснянському районах.
У птиці яєчного напрямку продуктивності ( ВАТ "Червоний прапор", СТОВ "Авіс", ВАТ "Сімейкінське") антитіла щодо адено - та реовірусів реєструвалися впродовж трьох років (2003-2005 рр.). При цьому титр антитіл коливався від 1,5 до 5,0 log2 у різні періоди вирощування. Антитіла виявляли фактично у 100% досліджених проб сироватки крові. Середні показники серопозитивної птиці з діагностичним титром (3,0 log2) по господарствах наведено на рисунках 1 та 2.
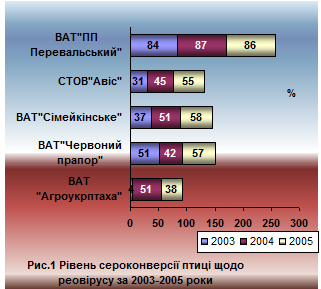

Оскільки вакцинопрофілактика проти рео - та аденовірусної інфекцій у господарствах яєчного напрямку продуктивності не проводилася, отримані результати серологічного моніторингу вказують на циркуляцію епізоотичних штамів рео - та аденовірусів. Необхідно зазначити, що антитіла до цих вірусів виявлялися протягом трьох років поспіль, а профілактичні засоби не проводилися проти жодної з інфекції, які вивчались, що свідчить про стаціонарність адено - та реовірусної інфекцій.
Для зменшення негативного впливу зазначених вірусів на рівень групового імунітету виникає необхідність введення до схем вакцинації птиці у господарствах щеплення проти реовірусної та аденовірусної інфекцій (АВІ) або корегування існуючих схем специфічної профілактики інфекційного бронхіту курей.
При аналізі ефективності вакцинопрофілактики ІБК на фоні циркуляції рео - та аденовірусів у трьох птахогосподарствах яєчного напрямку продуктивності найбільша ефективність щеплення проти ІБК встановлена у ВАТ "Червоний прапор". Використання 4-кратної імунізації птиці живими вакцинами різних штамів (а саме: 4/91 та Ма5) призвело до формування напруженого групового імунітету у 2004-2005р навіть на фоні циркуляції рео - та аденовірусів (рис. 3).
Використання схеми вакцинопрофілактики ІБК у 2003-2004 роках із застосуванням вакцини лише із штаму Ма5 у ВАТ "Сімейкінське" не забезпечувало формування напруженого імунітету до збудника ІБК. Проте, у 2005 році за умов застосування програми вакцинопрофілактики ІБК, що передбачала використання вакцин з різних штамів вірусу ІБК (4/91 та Ма5), груповий імунітет до ІБК на фоні циркуляції епізоотичних штамів рео - та аденовірусів у було сформовано на рівні 76%, що значно вище порівняно із 2003 (50%) та 2004 (62%) роками.
У СТОВ "Авіс" протягом 2003-2005 років птицю не щепили проти РВІ та АВІ, профілактика ІБК здійснювалась із застосуванням вакцини з одного штаму вірусу (Н-120). Групова імунна відповідь щодо ІБК була на невисокому рівні впродовж усього періоду спостереження (рис.3).
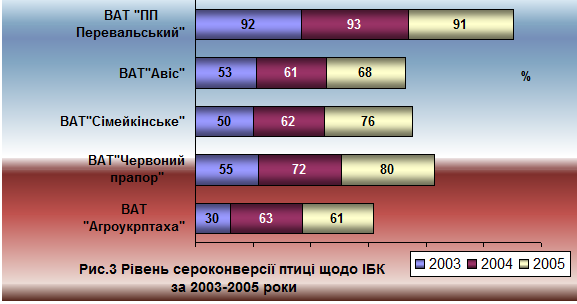
В господарстві мясного напрямку продуктивності ВАТ "Агроукрптаха" антитіла до аденовірусу 1 серотипу реєстрували у 100% дослідженої птиці на рівні від 3,0 до 5,0 log2. Титр антитіл до реовірусу коливався в межах від 1,0 - 3,0 log2 (2003 р.) до 3,0 - 4,0 log2 (2004 р.). Оскільки вакцинація проти РВІ не проводилась, наявність антитіл щодо цього збудника свідчить про циркуляцію епізоотичного штаму. У 2005 році в цьому господарстві впроваджено щеплення птиці проти РВІ, але з використанням лише однократної імунізації курчат у 10-добовому віці, що не призвело до формування напруженого імунітету до РВІ впродовж усього періоду вирощування та не вплинуло позитивно на формування імунітету до ІБК.
Систематичне щеплення птиці проти РВІ у ВАТ "ПП Перевальський" протягом трьох років за схемою дворазового застосування живої та одноразово - інактивованої вакцин забезпечувало формування напруженого імунітету під час імунізації птиці проти ІБК. Птиця весь період вирощування була захищена від інфікування епізоотичними штамами вірусу інфекційного бронхіту. Отже, профілактика РВІ за запропонованою нами схемою вакцинації, що передбачала триразове введення живої та однократне - інактивованої вакцин, позитивно впливало на формування імунної відповіді у птиці щодо вакцинних штамів ІБК.
Ізоляція, індикація та вивчення біологічних властивостей збудників реовірусної інфекції. За реовірусної інфекції, що встановлена нами серед птиці ВАТ "Агроукрптаха", хвора птиця відставала в рості та розвитку, в неї спостерігали артрити та теносиновіти. Схожі клінічні ознаки нами виявлено серед курчат-бройлерів, що утримувались у приватному секторі. Захворюваність сягала 80% як бройлерів, так і курей яєчного напрямку продуктивності, з патологічного матеріалу яких отримали ізоляти R-1 та R-2.
Під час ізоляції епізоотичних штамів реовірусу (R-1, R-2) в курячих зародках, виявлено застійні явища у внутрішніх органах, збільшення нирок, набряк ХАО. Типова ознака дїї реовірусів - утворення некротичних вогнищ на ХАО - в наших дослідженнях виявлена у 20% випадків у першому пасажі ізоляту R-1. Загибель ембріонів склала 20 та 40% у першому та другому пасажах (табл. 1).
Таблиця 1. Біологічна активніть ізолятів реовірусів птиці у курячих та гусячих зародках ( n=5)
|
Зміни |
У курячих зародках,% |
У гусячих зародках,% | ||||||
|
Ізолят R-1, пасаж |
Ізолят R-2, пасаж |
Ізолят R-1, пасаж |
Ізолят R-2, пасаж | |||||
|
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 | |
|
Набряк ХАО |
100 |
80 |
100 |
100 |
40 |
40 |
80 |
80 |
|
Вогнища некрозу на ХАО |
20 |
- |
- |
- |
- |
- |
20 |
- |
|
Затримка в розвитку |
20 |
40 |
- |
20 |
40 |
60 |
60 |
40 |
|
Некрози печінки |
- |
20 |
- |
- |
- |
20 |
40 |
- |
|
Застійні явища у внутрішніх органах |
100 |
100 |
100 |
100 |
- |
- |
- |
- |
|
Збільшення нирок |
60 |
80 |
100 |
80 |
20 |
40 |
100 |
60 |
|
Крововиливи по тілу зародка |
- |
- |
- |
- |
- |
20 |
- |
80 |
|
Загибель ембріонів |
20 |
40 |
20 |
20 |
- |
20 |
- |
- |
|
Примітка: - зміни не виявлені |
Розмноження ізолятів R-1 та R-2 у гусячих ембріонах характеризувалося затримкою в рості та розвитку ембріонів, крововиливами по тілу зародка, вогнищами некрозів на ХАО та печінки без застійних явищ у внутрішніх органах.
При визначенні інфекційної активності ізоляту R-1 у біопробі на курчатах встановлено, що захворюваність через тиждень складала 67%, через два тижні - 80%. Клінічні ознаки проявлялися вже на другу добу після інфікування курчат, в яких домінували проноси, збільшення гомілкових суглобів, слабкість кінцівок. Відмічено відставання в рості, зменшення на 25% маси інфікованих курчат через 7 діб після зараження порівняно з інтактними. За серологічними дослідженнями з реовірусним еритроцитарним антигеном виявили найбільший рівень титру антитіл (від 3,0 до 5,0 log2 ) через 10 діб після інфікування курчат.
Адаптація ізоляту R-1 до культивування у перещеплюваній культурі клітин VERO відбувалась уже з першого пасажу з розвитком цитопатогенної дії (ЦПД) через 120 годин після інфікування. Зі збільшенням кількості пасажів скорочувався термін прояву ЦПД вірусу. На четвертому пасажі округлення та утворення осередків уражених клітин моношару у вигляді синцитію виявляли вже через 22 години. Інфекційна активність ізоляту R-1 після четвертого пасажу становила 6,5 lg ТЦД50/см3(рис. 4).
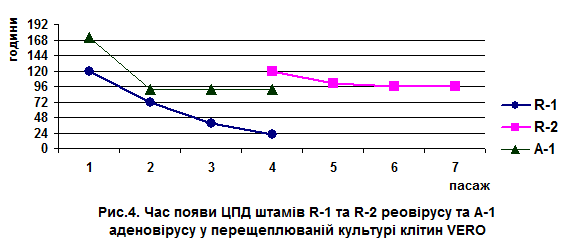
За результатами визначення інфекційності ізолят R-1 віднесено до високопатогенного, що підтверджується високим рівнем захворюваності курчат у біопробі з проявом типових клінічних ознак та ізоляції його в курячих та гусячих ембріонах вже з першого пасажу.
ЦПД ізолята R-2 реовірусу проявлялася лише в четвертому пасажі через 120 годин після інфікування моношару клітин. Після наступних трьох пасажів термін прояву ЦПД скорочувався на 24 години. Титр вірусу становив 5,0 lg ТЦД50/см3, що вказує на його менш виражені патогенні властивості в порівнянні з ізолятом R-1.
Ізоляція, індикація та вивчення біологічних властивостей збудників аденовірусної інфекції. Для ізоляції аденовірусів нами був використаний патологічний матеріал, відібраний у випадках прояву різних симптомокомплексів, які зустрічаються при аденовірусній інфекції курей. Так, ізолят А-1 був отриманий із матеріалу, відібраного від птиці із ознаками респіраторної інфекції, А-2 та А-3 - курячих ембріонів, що загинули на останніх етапах ембріогенезу, А-5 - завмерлого ембріону папуги Розели (Platycereus eximius), а ізолят А-6 - від птиці з ознаками гідроперикардиту.
Індикація ізолятів аденовірусу (А1, А-2, А-3, А-5) в курячих зародках характеризувалась відставанням у рості та розвитку ембріона, набряком підшкірної клітковини, гіпертрофією та утворенням локальних некрозів печінки, набряком та бляшками на ХАО.
У біопробі на курчатах ізоляту А-1 спостерігали пригнічення, зменшення апетиту, діарею. Смертність становила 17% інфікованого поголівя. За серологічними дослідженнями курчат виявили антитіла до аденовірусу 1 серотипу. При патологоанатомічному розтині загиблих курчат реєстрували ураження легень, печінки та нирок.
При культивуванні ізоляту аденовірусу А-1 у перещеплюваній культурі клітин VERO ЦПД проявлялась у вигляді округлення клітин через 172 години після інфікування у першому, та через 96 годин у 2-3 пасажах. Титр вірусу становив 4,0 lg ТЦД50/см3.
Ідентифікація ізолятів рео - та аденовірусів. Ідентичність ізолятів аденовірусу ( А-1, А-2, А-5, А-6) та реовірусу (R-1, R-2) визначалась за показниками електрофоретичних профілів вірусних білків у порівнянні з референтними штамами. Зокрема, поліпептиди IIIа та III пентону виявлено в ізоляті А-1. Поліпептид V серцевини та VIII гексону входив до складу ізолята А2 аденовірусу. В ізолятах А-5 та А-6 також знайдено білки з молекулярною масою, відповідною щодо маси основних структурних білків референтного штаму аденовірусу.
Результати досліджень електрофоретичних профілів польових ізолятів R-1, R-2 вказують на наявність основних білків реовірусу 1 та 2 в ізоляту R-1. В ізоляту R-2 виявлено основний 2 та мінорний 2 компоненти серцевини віріону, які є основною складовою референтного штаму реовірусу - СП-73.
Належність ізолятів А-1 та R-1, R-2 відповідно до адено - та реовірусів птиці підтверджено за допомогою реакції нейтралізації зі специфічними гіперімунними сироватками до референтних штамів аденовірусу (Phelps) та реовірусу ( СП-73) .
Результати реакції нейтралізації штамів R-1 та R-2 свідчать, що гіперімунна сироватка до референтного штаму СП-73 реовірусу нейтралізувала цитопатогенну дію ізолятів у перещеплюваній культурі клітин VERO. Титр ізоляту R-1 та R-2 під дією специфічної сироватки знизився на 3,0 lg ТЦД50/См3, що вказує на гомологічність досліджених ізолятів та гіперімунної сироватки, а відповідно і про ідентичність ізолятів R-1, R-2 та референтного штаму СП-73 реовірусу. Індекс нейтралізації ізолятів R-1 та R-2 специфічною гіперімунною сироваткою становив 1000 (табл.2).
Таблиця 2. Показники індексу нейтралізації специфічними гіперімунними сироватками референтних штамів рео - та аденовірусів
|
Індекс нейтралізації | |||
|
Штам |
Гіперімунна сироватка крові до штаму СП-73 реовірусу |
Штам |
Гіперімунна сироватка крові до штаму Phelps аденовірусу |
|
СП-73 |
1000 |
Штаму - Phelps |
1000 |
|
R-1 |
1000 |
Штам А-1 |
100 |
|
R-2 |
1000 |
Ідентичність ізоляту А-1 референтному штаму Phelps аденовірусу птиці встановлена за нейтралізацією специфічною гіперімунною сироваткою до штаму Phelps у перещеплюваній культурі клітин VERO. При цьому інфекційний титр вірусу знизився на 2,0 lg ТЦД50/См3, що на 1,0 lg менше, ніж за нейтралізації штаму Phelps цією ж сироваткою. Індекс нейтралізації ізоляту А-1 специфічною гіперімунною сироваткою становив 100.
Експериментальне визначення впливу рео - та аденовірусів на напруженість імунітету щодо вакцинного штаму вірусу інфекційного бронхіту. Для досліду щодо визначення впливу рео - та аденовірусів на ефективність вакцинопрофілактики ІБК було сформовано 7 груп курчат, які були попередньо інфіковані епізоотичними та референтними штамами рео - та аденовірусів, через 5 тижнів після цього - щеплені проти ІБК живою вакциною (штам Н-120). Серологічні дослідження сироваток крові курчат проводили через 2 та 5 тижнів після вакцинації курчат.
Встановлено достовірну різницю у рівні імунної відповіді до вірусу ІБК курчат, інфікованих різними штамами реовірусу птиці, та контрольних. Так, в інфікованих піддослідних 1 і 2 груп курчат як польовим (R-1), так і референтним (СП-73) штамами реовірусу титр післявакцинальних антитіл після щеплення вакциною проти ІБК достовірного знаходився на рівні 3 ±0,31 log2 та 3±0,32 log2, відповідно, в той час як у контрольній групі імунізованих проти ІБК інтактних курчат рівень антитіл досягав 5±0,31 log2 через 2 тижні після вакцинації (табл. 3).
Вірус інфекційний бронхіт птиця
Таблиця 3. Рівень антитіл до рео - аденовірусу та вірусу ІБК у курчат дослідних груп (n=5)
|
Групи курчат |
Штами вірусів |
Рівень антитіл щодо вірусів у РНГА, log2 | |||||||
|
Через 3тижні п/з |
Через 7 тижнів п/з, Через 2 тижні п/в |
Через 10 тижнів п/з, Через 5 тижнів п/в | |||||||
|
Рео Вірус |
Адено Вірус |
Рео Вірус |
Адено Вірус |
Вірус ІБК |
Рео Вірус |
Аденовірус |
Вірус ІБК | ||
|
1 |
R-1 |
2,2 ±0,37 |
- |
|
- |
|
|
- |
2,9±0,4*** |
|
2 |
СП-73 |
2±0,45 |
- |
|
- |
|
|
- |
2,8±0,36*** |
|
3 |
А-1 |
- |
2±0,31 |
- |
|
|
- |
|
3,5±0,31** |
|
4 |
Phelps |
- |
|
- |
|
|
- |
|
3,5±0,26** |
|
5 |
Phelps + СП-73 |
2,6±0,24*** |
2±0,31 |
|
|
|
|
|
|
|
6 |
А1+ R-1 |
1±0,32 |
|
4±0,31 |
3±0,32 |
** |
|
|
*** |
|
7 |
Контрольна |
- |
- |
- |
- |
5±0,31 |
- |
- |
5±0,32 |
|
N=кількість голів у групі; п/з - після зараження ; п/в - після вакцинації ***Р<0,001 ;** Р<0,01; ** Р<0,05 дослідні групи відповідно контрольних |
Інфікування курчат аденовірусами також пригнічувало імунну відповідь щодо вірусу ІБК, але в меншій мірі. Через два тижні після вакцинації антитіла до вірусу ІБК реєстрували на рівні 3,8±0,2 Log2 та 3,4±0,4 Log2 у курчат, інфікованих ізолятом А-1 та штамом Phelps, відповідно, що на 1,2 та 1,6 Log2 менше, ніж у контрольній групі.
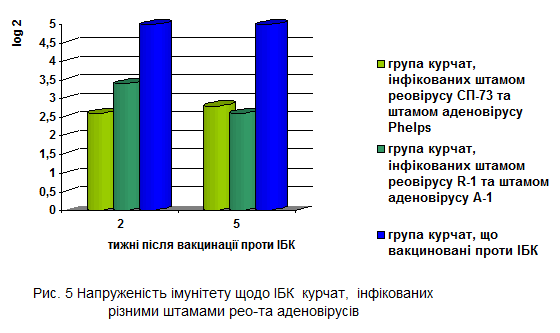
Менший рівень антитіл щодо ВІБ майже в два рази зареєстровано через 2 тижні після вакцинації у курчат 5-ї групи (інфікованих референтними штамами СП-73 рео - та Phelps аденовірусів) порівняно з контрольною групою курчат.
Найбільшу імуносупресивну дію епізоотичних ізолятів R-1 та А-1 встановлено через 5 тижнів після вакцинації курчат проти ІБК, вірусоспецифічні антитіла до якого реєстрували лише на рівні 2,6±0,24 log2, що достовірно відрізнялося від показників у курчат контрольної групи (рис.5).
Імуносупресивні властивості рео - та аденовірусів підтверджено результатами гістологічних досліджень основного імунокомпетентного органу птиці - бурси Фабриціуса. У курчат, інфікованих референтними штамами та польовими ізолятами рео - та аденовірусів (групи курчат 1, 2, 3, 4), спостерігали різний ступінь деструктивних змін, а саме: гіпоплазію фолікулів, зменшення їх кількості та залозисте переродження, місцями заміщення фолікулів сполучною тканиною. Польовий ізолят аденовірусу А-1 також обумовлював руйнування епітелію слизової оболонки бурси Фабриціуса.
У курчат, одночасно інфікованих рео - і аденовірусами (групи 5 та 6), спостерігали найбільш тяжкі зміни бурси Фабриціуса: близько 50% фолікулів були частково делімфатизовані і не мали коркової речовини. Розмір фолікулів, які були розділені широкими прошарками сполучної тканини, складав 100-160 мкм. Значна частина органу представлена залозистими структурами, які утворилися на місці делімфатизованих фолікулів.
Отже, шляхом використання серологічних та гістоморфологічних методів підтверджено супресивну дію рео - і аденовірусів на органи імунної системи курчат. В той же час моноінфекція реовірусу спричиняє більш виражений імуносупресивний стан в організмі птиці, ніж аденовіруси. В асоціації рео - та аденовіруси посилюють імуносупресивні властивості один одного.
Похожие статьи
-
Роботу виконано в Науково-виробничому центрі ветеринарної медицини птахівництва Луганського національного аграрного університету. Окремі дослідження...
-
Актуальність теми. На сьогодні в промисловому птахівництві широко застосовується щеплення птиці проти основного особливо небезпечного вірусного...
-
Антисиро-ватки до Польових І референтних штамів Нейсерій Титри антитіл з антигенами N. spp. turkies N. spp. hens N. spp. ducks N. ovis N. canis N. caviae...
-
Епізоотологічний моніторинг щодо нейсеріозу птиці в регіонах України. З завозом імпортної птиці в птахогосподарствах багатьох регіонів країни почали...
-
Вплив КБП і ПМД на розвиток збудника на вегетуючих рослинах. При дослідженні застосування біологічних препаратів БСП, КБП, полімінерального добрива як...
-
1. Вперше в Україні проведено епізоотологічний моніторинг, визначено поширення та етіологічну роль бактерій Neisseria spp. у патології...
-
№ П/п Назва господарств, Де проводили Виробничу перевірку Рік Досліджень Вид Птиці Всього Поголів'я Кількість вибракованих голів (в міс.) До Після %...
-
Клініко-епізоотологічні дослідження. При обстеженні з допомогою епізоотологічного, клінічного і лабораторних методів стада зі 130 корів одного...
-
№ П/п Область Вид птиці Рік Досліджен-ня Вік птиці (діб) Виявлено клінічно хворих на період дослідження (%) 1 Харківська Індики Качки Кури 1996 1994...
-
Ріст та розвиток сої Висота рослин у період вегетації є важливим загальним показником. За цим показником можна побачити рівень забезпеченості рослин...
-
N П / П Вид птиці, Порода, крос Вік, дні Кіль-кість Проб Титри антитіл Log2 М±м 1 Качки: "Темп" "Благоварський" Українська біла Дикі - "кряква" 360...
-
Робота виконана впродовж 1990-2003 років у відділі профілактики хвороб птиці Інституту птахівництва УААН. Дослідження проводили на індиках кросу "Хідон"...
-
Способи зближення цвітіння батьківських форм гібридів кукурудзи У середньому за 2002-2005рр. періоди "сходи-цвітіння качанів" та "сходи-повна стиглість"...
-
Проведено аналіз літературних джерел з питань впливу способів основного обробітку грунту та добрив на агрохімічні, агрофізичні, мікробіологічні...
-
Динаміка росту дослідних груп тварин. Вивчався ріст молодняку на всіх етапах постнатального розвитку, починаючи від дня народження до 18 місяців. Для...
-
Поширеність та причини виникнення асептичного пододерматиту у великої рогатої худоби За результатами хірургічної диспансеризації поголів'я великої...
-
Целью данной курсовой работы является углубление, закрепление и практическое применение знаний, полученных в процессе изучения курса...
-
Продуктивный потенциал птицы во многом закладывается во время выращивания, поэтому этот период чрезвычайно важен. При этом решающее значение имеет живая...
-
Виявлено, що у корів, які в зимово-стійловий період перебували на залізобетонній щілинній підлозі, на 9,1 та 8,5% знижувався показник вологи копитцевого...
-
Бройлеров выращивают во всех регионах мира, в условиях широкого диапазона температур и относительной важности. Вследствие такой вариации зоотехникам,...
-
Правила отбора яиц для инкубации. Для инкубации следует отбирать яйца средней массы, а масса яйца зависит от породы и возраста птицы, а также от ее...
-
В первые 14 дней жизни цыпленка закладывается фундамент его успешного развития и роста. Каждый дополнительный грамм живой массы в возрасте семи дней...
-
Успех выращивания бройлеров существенно зависит от правильной плотности посадки, что обеспечивает эффективное использование площадей для получения...
-
Работа с подстилкой становится еще одним важным аспектом управления микроклиматом. Хорошее состояние подстилки является одной из предпосылок для здоровья...
-
Для достижения оптимальных результатов при выращивании бройлеров важно, чтобы микроклимат в птичнике отвечал потребностям птицы. Для этого необходимо...
-
Одним из ключевых элементов для достижения максимальных результатов при выращивании птицы является обеспечение устойчивого микроклимата в птичнике....
-
У дослідженнях різних способів зближення строків цвітіння качанів пізньостиглої інбредної лінії Гк 26 і волотей ранньостиглої Чк 73 економічно...
-
В птичнике для содержания ремонтного молодняка должны быть следующие вспомогательные помещения: Помещение для обслуживающего персонала. Служит местом...
-
Независимо от применяемого типа системы кормления, фронт кормления является наиболее важным фактором. Если фронт кормления недостаточен, то скорость...
-
Наливайко Л. І. Нейсеріоз сільськогосподарської птиці (епізоотологічний моніторинг, діагностика та засоби профілактики). - Рукопис. Дисертація на...
-
Програма досліджень передбачала: Визначення динаміки висоти рослин сої : 4 рази. Висоту рослин вимірювали рулеткою від поверхні грунту до верхівки...
-
(огляд літератури) Проведено аналіз результатів досліджень вітчизняних і закордонних вчених в області біології культури гречки та технології її...
-
Эмбриональное развитие птиц возможно лишь при определенных внешних условиях. Необходимы соответствующий обогрев яиц, достаточно влажный, чистый и...
-
Польові дослідження виконували на Носівській селекційно-дослідній станції Чернігівського ІАПВ УААН у тимчасових дослідах протягом 2002-2004 рр. Грунт...
-
Интерьер птицы и методы его изучения - Конституция, экстерьер и интерьер сельскохозяйственной птицы
Интерьер - совокупность внутренних физиологических, анатомо-гистологических и биохимических свойств организма; находится в тесной взаимосвязи с...
-
Экстерьер птицы и методы его оценки - Конституция, экстерьер и интерьер сельскохозяйственной птицы
Экстерьер, или внешние формы и признаки строения тела сельскохозяйственной птицы, связан с ее конституцией и хозяйственно-полезными качествами. По данным...
-
Как распознать и лечить колибактериоз? - Бактериальные болезни птиц
Основная цель данной статьи, чтобы помочь человеку, невооруженному специальными знаниями, распознать болезнь и предпринимать первые шаги по ее лечению....
-
Актуальність теми. Птахівництво - одна з галузей тваринництва, яка першою стала на шлях інтенсивного розвитку і за короткий період вийшла на передові...
-
КОКЦИАИОЦИДНОЕ и кокцидиостатическое действие. - Бактериальные болезни птиц
Некоторые лекарственные препараты ( Кокцидиоцидные ) уничтожают паразита, а другие ( Кокцидиостатические ) только сдерживают его развитие. После...
-
ЭПИЗООТОЛОГИЯ - Бактериальные болезни птиц
Куры являются единственным естественным хозяином для видов кокцидий, описанных выше. Сообщения об инвазировании этими видами Eimeria других птиц не...
РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ - Рео - та аденовірусна інфекції і їх вплив на ефективність вакцинопрофілактики інфекційного бронхіту птиці