ОБЗОР ЛИТЕРАТУРЫ ПО ТЕМЕ ИССЛЕДОВАНИЯ, Физиологические основы атлетической гимнастики - Основы периодизации круглогодичной тренировки
Физиологические основы атлетической гимнастики
В организме человека различают Скелетные, гладкие мышцы и сердечную мышцу.
Гладкие мышцы входят в состав внутренних органов тела человека.
Сердечная мышца занимает промежуточное положение по своим функциональным свойствам между скелетными и гладкими мышцами: она трудно управляется волевыми усилиями, но имеет чрезвычайно высокую работоспособность. Как и скелетная мышца, она может сокращаться быстро и мощно, но в то же время долго работать.
Скелетные мышцы являются активными элементами двигательного аппарата человека. Они образованы поперечно-полосатыми мышечными волокнами. Каждое мышечное волокно окружено прозрачной оболочкой, содержащей эластичные Коллагеновые нити. Небольшие группы мышечных волокон заключаются в оболочку из соединительной ткани (эндомизий), а более крупные пучки мышечных волокон и мышца в целом окружены рыхлой соединительной тканью. Все соединительные мышечные структуры непрерывно связаны между собой и являются продолжением друг друга. Они образуют параллельный эластический элемент мышцы.
У большинства скелетных мышц для удобства описания различают Брюшко и Два конца. Один из этих концов является началом мышцы и называется ее Головкой, а противоположный конец получил название Хвоста Мышцы (рис. 1). У концов мышцы соединительная ткань образует сухожилия, которыми мышца прикрепляется к костям скелета. Эти сухожилия образованы пучками коллагеновых волокон, параллельно вытянутых по длине мышцы. Некоторые относительно плоские мышцы имеют соответственно и плоские сухожилия - Апоневрозы (рис. 1).
Сверху каждая мышца покрыта оболочкой, которая называется Фасцией. Фасции представляют собой пластины из соединительной ткани с большим количеством коллагеновых и эластических волокон, имеющих различную протяженность, толщину и количество слоев. Ориентация этих волокон по отношению к продольной оси мышцы обусловлена функциональными особенностями каждой мышцы или группы мышц, покрытых данной фасцией. Фасции могут располагаться и между мышцами в виде перегородок, или срастаться с надкостницей, образуя влагалища, к стенкам которых прикрепляются мышцы. В наиболее подвижных частях скелета, например, в области кисти или стопы, имеются волокнистые сухожильные влагалища, которые облегчают скольжение сухожилий в строго определенных направления. По расположению мышечных волокон и их отношению к сухожилию различают три основных типа скелетных мышц.
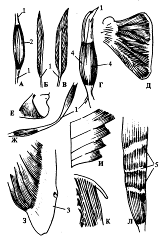
Рис. 1. Формы и строение скелетных мышц: А - веретенообразная мышца; Б - одноперистая м.; В - двуперистая м.; Г - двуглавая м.; Д - многоперистая м.; Е - треугольная м.; Ж - двубрюшная м.; 3 - широкая мышца, имеющая апоневроз; И - зубчатая м.; К - квадратная м.;Л - мышца с сухожильными перемычками: 1 - сухожилие; 2 - брюшко мышцы; 3 - апоневроз; 4 - головка мышцы; 5 - сухожильная перемычка (по Синельникову Р. Д., Синельникову Я. Р., 1989).
- 1. Параллельный, у которого пучки мышечных волокон расположены параллельно продольной оси мышцы. Одни мышцы этого типа могут иметь от одной до четырех головок, а другие - два брюшка, разделенных сухожильными перемычками (рис. 1 - А, Г, Ж). 2. Перистый, в котором параллельно расположенные пучки мышечных волокон находятся под углом к продольной оси мышцы. Этот тип мышц может иметь несколько форм. Различают одноперистые мышцы, пучки волокон которых располагаются по одну сторону сухожилия, двуперистые - по обе стороны сухожилия, а также многоперистые мышцы, пучки волокон которых примыкают друг к другу несколькими перистыми сегментами (рис. 1-Б, В, Д). 3. Треугольный тип - когда мышечные пучки сходятся с различных направлений к одному общему хвосту (рис. 1-Е).
К каждой мышце подходят нервы, управляющие ее работой, и сосуды, снабжающие мышцу кровью.
Целенаправленная Физическая тренировка оказывает положительное влияние не только на скелетные мышцы. Благодаря, ей улучшается функциональное состояние и гладкой мускулатуры, и сердечной мышцы. Например, тренировкой на выносливость можно увеличить массу сердечной мышцы и повысить эффективность ее работы, что, в свою очередь, приводит к повышению работоспособности человека. А хорошо развитый "мышечный корсет" - не только признак силовой подготовленности: он также создает благоприятные условия и для деятельности внутренних органов, способствует улучшению работы пищеварительной системы. В конечном итоге, все это приводит к улучшению энергетического обеспечения мышечной деятельности и укреплению здоровья.
Физическая тренировка оказывает благотворное влияние на весь организм человека, на все виды мышечной ткани и на все системы жизнеобеспечения. При этом особая роль в осуществлении двигательной активности человека принадлежит скелетной мускулатуре.
Скелетные мышцы составляют активную часть двигательного аппарата, являясь преобразователями химической энергии непосредственно в механическую работу и тепло.
Каждая скелетная Мышца представляет собой орган, состоящий из мышечных Клеток-миофибрилл, соединительной ткани, сосудов, нервов (рис.2).
Один грамм мышечной ткани содержит примерно 100 мг сократительных белков Актина И миозина. Эти белки образуют в способных к сокращению Миофибриллах тонкие и толстые нити, которые располагаются параллельно вдоль мышечной клетки. В микроскоп можно увидеть В миофибриллах чередующиеся темные и светлые поперечные Полосы, из-за чего скелетные мышцы и получили название поперечно-полосатых. Эта поперечная исчерченность обусловлена особой регулярной организацией нитей Актина и Миозина. Поперечные темные перегородки, названные Z-пластинками, разделяют Миофибриллы на Саркомеры - структурно-функциональные единицы сократительного аппарата. В середине каждого Саркомера расположены несколько тысяч "толстых" Миозиновых Нитей, а на его обоих концах - до 2000 "тонких" Актиновых нитей, прикрепляющихся к Z-пластинкам наподобие щетинок в щетке. Оптическая неоднородность Саркомеров позволяет выделить в каждом из них Светлые 1-диски, более Темные А-диски, а также центральную Н-зону. 1-диски тянутся до Z-пластинок. В покоящихся мышцах I - и А - диски незначительно перекрываются, и, поэтому, в микроскоп кажутся несколько темнее, чем центральная Н-зона, в которой нет Актиновых Нитей. На электронных микрофотографиях можно также обнаружить в H-зоне центральную М-линию, определяемую как сеть опорных белков, удерживающих вместе в виде пучка толстые нити миозина в середине саркомера (рис. 2).
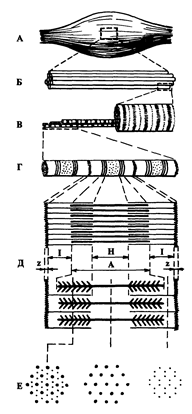
Рис. 2. Схематическое изображение строения мышцы. А - мышца; Б - пучок мышечных волокон; В - мышечное волокно; Г - миофибрилла; Д - схематическое изображение саркомера, ограничено Z-линиями, с 1-дисками, А-дисками и Н-зоной; Е - поперечный срез на различных участках саркомера, дающий представление о распределении тонких и толстых нитей актина и миозина (по Я. Коцу).
Мышца сокращается благодаря скольжению тонких Актиновых нитей вдоль толстых нитей Миозина, двигаясь между ними к середине Саркомера. Сами Актиновые и Миозиновые нити не укорачиваются, их длина не изменяется и при растяжении мышечной ткани: лишь пучки тонких Актиновых нитей, скользя между толстыми нитями Миозина, Выходят из промежутков между ними так, что степень их взаимного перекрытия может уменьшиться до нуля (рис. 2).
Разнонаправленное скольжение Актиновых и Миозиновых нитей в соседних половинках Саркомеров осуществляется за счет того, что нити миозина имеют поперечные выступы, называемые "мостиками", или Головками миозина (рис. 3). Каждый такой поперечный "мостик" во время сокращения и Связывает миозиновую нить с Актиновой: "наклоны" головок создают объединенное усилие и выполняют как бы "гребок", продвигающий нить Актина к середине Саркомера. При однократном движении поперечных мостиков вдоль Актиновой нити Саркомер может укоротиться только примерно на 1 % своей длины. Однако, мышцы при сокращении могут укорачиваться до 50% своей длины. При этом поперечные мостики делают не один "гребок", а как бы выполняют серию гребковых движений не один, а 50 раз за то же время. Благодаря суммации таких ритмических "гребков" и соответствующих им укорочений последовательно расположенных в Миофибриллах саркомеров, мышца может развить большую силу.
Ритмические повороты Головок миозина при этом "гребут" Актиновые Нити к середине Саркомеров. Этот процесс для образности сравнивают с группой - людей, тянущих длинные веревки, перебирая их руками.
Вместе с тем, Саркомер может генерировать силу и без относительного скольжения нитей, то есть без изменения своей длины - в строго изометрических (статических) условиях. Это является возможным благодаря эластичности поперечных "мостиков". Так как каждое мышечное волокно состоит из большого количества последовательно расположенных Саркомеров, то величина силы тяги, развиваемая этим волокном, и скорость его укорочения возрастают во столько раз, во сколько длина Миофибриллы Больше длины составляющих ее Саркомеров. Поэтому более длинная мышца будет сокращаться сильнее и быстрее, чем короткая. Например, портняжная мышца лягушки сокращается всего лишь со скоростью 0,2м/сек, причем каждый ее Саркомер из исходной длины 2 мкм за 50 мс укорачивается до длины 1 мкм. А мышцы руки человека, имея гораздо большую длину, укорачиваются уже со скоростью 8,0 м/сек.
Чем больше Миофибрилл будет в мышце, тем она будет сильнее. Поэтому проявляемая мышцей сила пропорциональна ее физиологическому поперечнику.
Мышечные волокна функционально объединены в Двигательные единицы (ДЕ). ДЕ состоят из одного Мотонейрона и группы иннервируемых им мышечных волокон (рис. 4). Состав различных мышц человека различается по количеству ДЕ. Значительно варьируют и размеры ДЕ - один Мотонейрон может иннервировать от нескольких мышечных волокон до 500 - 2000. Количество волокон в ДЕ одной и той же мышцы также не одинаково. Каждое мышечное волокно состоит Из миофибрилл (рис. 4).
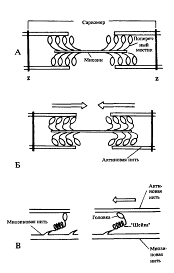
Рис. 3. Схематическое изображение поперечных мостиков саркомера. Миозиновая нить с поперечными мостиками, соединенная с актиновыми нитями, А - до, Б - во время "гребковых движений" (эти движения происходят асинхронно), В - модель механизма генерации силы поперечными мостиками до (слева) и во время (справа) "гребкового движения" (по Ruegg J. С., 1983).
Мышцы, выполняющие "тонкую" и точную работу, например, мышцы глаз, пальцев рук и т. п, обладают большим количеством ДЕ (от 1500 до 3000), но состоят эти ДЕ из малого количества Миофибрилл - от 8 до 50. В противоположность им, мышцы рук, ног или спины, выполняющие относительно более "грубые" и менее точные движения, но требующие большой силы, имеют гораздо меньшее количество ДЕ, состоящих из большого числа мышечных волокон: от 600 до 2000. Например, ДЕ Бицепса плеча могут включать 1000, 1200, 1400 или даже 1600 миофибрилл. В ДЕ Большеберцовой мышцы их около 1600, а в Мышцах спины количество иннервируемых одной ДЕ Миофибрилл достигает 2000.
ДЕ состоят из двух основных типов мышечных волокон:
Быстрых и сильных, но быстро утомляемых FT-волокон (Fast - быстрый, Twitch - сокращение);
Выносливых, но менее сильных и быстрых ST-волокон (Slow - медленный).
Быстрые мышечные волокна имеют повышенное содержание Гликогена, высокую активность Анаэробных гликолитических ферментов, обеспечивающих использование внутримышечных Энергетических субстратов, а потому они менее приспособлены для длительной работы, обеспечиваемой преимущественно Аэробным (окислительным) способом энергопродукции. Не обладая большой выносливостью, эти волокна наиболее приспособлены для быстрых и сильных, но относительно кратковременных мышечных сокращений, обеспечивая выполнение кратковременной физической работы высокой мощности продолжительностью не более 4-х минут (к примеру - бег на короткие или средние дистанции). По международной номенклатуре быстрые мышечные волокна еще обозначаются как FG-тип (быстрые, Glycolysis - гликолигические).
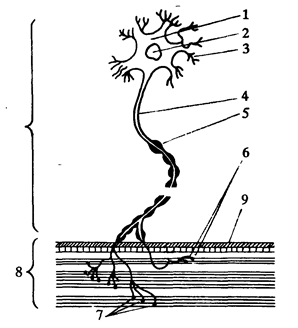
Рис. 4. Схема строения двигательной единицы. 1 - тело мотонейрона; 2 - ядро; 3 - дендриты - короткие отростки; 4 - аксон - длинный отросток нервного волокна; 5 - оболочка нервного волокна, 6 - концевые веточки аксона; 7 - нервно-мышечные синапсы, через которые мышце передаются нервные импульсы; 8 - мышечное волокно с миофибриллами; 9 - оболочка мышечного волокна (по Я. Коцу).
Медленные мышечные волокна более приспособлены для обеспечения длительных, но менее мощных по силе мышечных сокращений, что характерно для выполнения продолжительной работы на выносливость. Медленные волокна имеют широко разветвленную Капиллярную сеть, что позволяет им получать большое количество кислорода из крови. Эти волокна отличаются также повышенным содержанием Миоглобина и наличием большого количества Митохондрий (внутриклеточных структур, в которых протекают процессы окисления), характеризуются высокой активностью Окислительных ферментов и имеют более высокое содержание жиров в виде Триглицеридов - субстратов окисления. По международной номенклатуре медленные волокна обозначаются как SO-тип (Slow - медленные, Oxydative - окислительные).
Вместе с тем, среди быстрых волокон выделяют подтип быстрых Окислительно - Гликолитических, по международной терминологии fog-тип. Эти волокна приспособлены к достаточно интенсивной Окислительной (аэробной) работе с одновременным мощным Гликолитическим энергообразованием, однако, их окислительные возможности ниже, чем у медленных окислительных волокон. С функциональной точки зрения они рассматриваются Как промежуточный тип между двумя основными FG - и SO-типами мышечных волокон.
Композиционный состав мышц определен Генетически: в течение жизни общее количество и соотношение имеющихся в мышцах типов волокон не изменяется. Под воздействием тренировки может изменяться толщина волокон всех типов, а значит, способность мышц к выполнению физической работы различной физиологической направленности.
У нетренированных молодых мужчин наибольшую толщину имеют волокна промежуточного FOG-типа, у молодых женщин - SO-типа. У мужчин в возрасте 16-30 лет, благодаря высокой двигательной активности, увеличивается толщина всех трех типов мышечных волокон. Визуально это выражается в наращивании, у юношей объема мышечной массы, "возмужании". У женщин в этом периоде жизни мышечные волокна, наоборот, имеют тенденцию к утончению. Специалисты связывают это со снижением женщинами в этом возрасте интенсивности мышечной деятельности и двигательной активности в целом. Целенаправленными тренировочными занятиями можно существенно изменить общий объем мышечной массы различных сегментов тела и функциональные возможности мышц. Например, у некоторых тяжелоатлетов площадь поперечного среза Четырехглавой мышцы бедра до 90% может состоять, из быстрых и сильных FG-волокон, а у марафонцев - из медленных, но выносливых SO-волокон.
Управление движениями, поддержание вертикального положения и необходимая фиксация звеньев тела обеспечивают сокращение в нужный момент времени определенных мышц и регуляцией степени их напряжения Центральной нервной системой.
Регуляция мышечного напряжения осуществляется тремя физиологическими механизмами:
- 1) количеством активных ДЕ мышцы; 2) частотой импульсации Мотонейронов ДЕ (т. е. режимом их работы); 3) временной связью активности ДЕ. ДЕ активизируется после того, как ее Мотонейрон пошлет импульсы для сокращения иннервируемых мышечных волокон.
Двигательные единицы возбуждаются Мотонейронами по физиологическому закону "все или ничего". Поэтому на нервный импульс реагируют одновременно все мышечные волокна одной ДЕ. Сила сокращения одной ДЕ зависит от количества составляющих ее мышечных волокон. Малые ДЕ развивают силу всего лишь в. несколько Миллиньютон, а ДЕ с большим количеством волокон - в несколько Ньютон. Как видно, силовой потенциал одной ДЕ невелик, поэтому для выполнения движения при сокращении мышцы одновременно "включаются" в работу несколько ДЕ, что в физиологии получило название "пространственной суммации". Чем выше внешнее сопротивление, тем больше ДЕ задействовано при генерации силы мышцей, и тем большее напряжение она развивает.
Необходимое число активных ДЕ определяется интенсивностью возбуждающих влияний более высоких уровней нервной системы на Мотонейроны данной мышцы.(3)
Реакция Мотонейронов ДЕ на возбуждающие влияния более высоких уровней нервной системы определяется порогом их возбуждения. Сам этот Порог зависит от размера Мотонейрона. Чем меньше размер тела Мотонейрона, тем ниже порог его возбуждения и меньше размер ДЕ. Поэтому слабые мышечные напряжения обеспечиваются преимущественно активностью низко пороговых - малых и медленных ДЕ.
В естественных условиях сокращение ДЕ работающих мышц обусловлено не одиночными нервными импульсами, а их сериями с различной частотой - от 5 до 50 в одну секунду. При этом, когда каждый последующий нервный импульс подается до окончания фазы расслабления мышцы от воздействия предыдущего, то последующее за импульсом сокращение мышцы накладывается на предыдущее. В итоге происходит более высокое развитие силы. Когда нервные импульсы генерируются Мотонейроном с высокой частотой, то спада напряжения мышц или развиваемой ими силы не происходит. При этом достигаются более сильные, чем при одиночных импульсах, сокращения мышечных волокон и 3-4-кратное увеличение развиваемой силы. Такое сокращение мышц называется Титаническим (рис. 5). Продолжительность Титанического сокращения ДЕ может в десятки и даже в тысячи раз превышать продолжительность ее сокращения от одиночного нервного импульса.
Если постепенно увеличивать напряжение мышц, то в работу будут вовлекаться все более крупные по размеру ДЕ: начиная от малых низко пороговых и до больших - высоко-пороговых.
Выносливые SO-волокна обладают низким порогом возбуждения. Частоты в 20 импульсов в секунду (20 Гц) уже может быть достаточно для полного исчерпания их силового потенциала.
В связи с тем, что FG-волокна сокращаются и расслабляются гораздо быстрее, чем SO-волокна, Частота импульсации для достижения их максимальной активации должна быть выше. Поэтому при частоте 25-30 Гц достигается лишь умеренное по силе напряжение у этого типа волокон. Максимальное напряжение и максимальная сила достигаются у них лишь при частотах 45-50 Гц. Мышца, состоящая преимущественно из ДЕ с SO-во-локнами, может поддерживать максимальный уровень своего напряжения значительно дольше, чем состоящая преимущественно из ДЕ с FG-волокнами. Но развиваемая при этом сила, очевидно, будет существенно ниже.
В связи с этим, необходимая для полного Титанического сокращения ДЕ частота импульсации Мотонейрона зависит и от типа составляющих ее мышечных волокон.
Регулировка силы сокращения мышц за счет изменения частоты нервной импульсации определяется термином "временная суммация".
Когда скелетные мышцы преодолевают легкие и умеренные сопротивления, например, в быту или в процессе обычной трудовой деятельности, необходимая для их возбуждения частота нервной импульсации не достигает максимума, а деятельность низко пороговых ДЕ при этом осуществляется попеременно, т. е. асинхронно. В этом случае относительно низкое общее напряжение мышцы не колеблется, поскольку для разных асинхронно активных ДЕ максимумы сокращений никогда не совпадают.
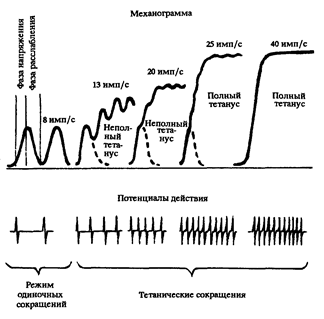
Рис. 5. Режимы сокращений двигательной единицы импульсами различной частоты (по Я. Коцу).
При длительной мышечной работе умеренной интенсивность, например, в длительном беге или лыжных гонках, в первую очередь активизируются низко пороговые ДЕ. По мере утомления этих работающих ДЕ, их сократительная способность постепенно уменьшается, и в работу начинают вовлекаться более крупные и высоко пороговые ДЕ. Этим объясняется тот факт, что в процессе длительной работы умеренной интенсивности Гликоген - внутримышечный источник энергии - более быстро расходуется в медленных SO-волокнах, а по мере продолжения работы и в быстрых FG-волокнах.
При скоростном проявлении силы и необходимости при этом преодоления мышцами умеренных и высоких внешних сопротивлений, происходит активизация преимущественно высоко пороговых ДЕ с FG-волокнами короткими сериями высокочастотных нервных импульсов. Такая Стартовая иннервация Вызывает сильный и нарастающий процесс мышечного сокращения, после чего за "взрывным" началом следует Сигнальная блокировка биоэлектрической активности мышц. Во время этой "паузы" ДЕ и сокращаются с высокой скоростью. Движения при таком баллистическом характере мышечного сокращения заранее программируются в головном мозге и реализуются с такой высокой скоростью, что их коррекция по ходу движения часто оказывается практически невозможной. Период Биоэлектрического "молчания", во время которого мышца не реагирует на поступающие к ней нервные импульсы, следующие за Стартовой иннервацией, зависит преимущественно от величины преодолеваемого внешнего сопротивления.
Если внешнее сопротивление возрастает настолько, что при Баллистическом сокращении мышц ускорения движения не происходит, то подается новая серия импульсов с последующим периодом "молчания", что в конечном итоге до определенного уровня величины внешнего сопротивления и обеспечивает дальнейшее ускорение выполняемого движения.
Движения, характеризуемые сериями импульсов Стартовой иннервации с последующим периодом Сигнальной блокировки и проявлением Баллистического режима мышечного сокращения, имеют резко выраженный Скоростно-силовой характер.
Если же сопротивление увеличивается до максимального или близкого к нему, то такое сопротивление будет преодолеваться уже более продолжительными сериями импульсов очень высокой частоты. Такой режим работы мышц характерен для проявления Максимальной силы.
Механические свойства мышц достаточно сложны и зависят от свойств образующих их компонентов: Миофибрилл, соединительных и Трофических образований и т, п., а также от состояния самой мышцы.
Для понимания большинства свойств мышечной механики можно использовать упрощенную модель ее строения. Эта модель представляет собой комбинацию сократительных и упругих компонентов мышцы (рис. 6).
Сократительные элементы соответствуют участкам Саркомеров, где Актиновые и Миозиновые нити перекрывают друг друга. В этих участках при возбуждении мышцы и происходит генерация мышечной силы.
Актиновые и Миозиновые нити легко скользят относительно друг друга, поэтому в расслабленных мышцах сопротивление растяжению оказывается очень низким. Укороченную при сокращении мышцу небольшим усилием можно снова растянуть до исходной длины. Однако это растяжение мышцы до своей исходной длины является пассивным процессом, который может быть осуществлен лишь за счет внешнего воздействия.
Если же покоящуюся мышцу потянуть за один конец так, чтобы ее волокна растянулись, то после снятия внешней нагрузки мышца восстанавливает свою длину. Это говорит о том, что в ней развивается Пассивное напряжение. Значит, покоящаяся мышца обладает эластичностью, которая свойственна преимущественно Растяжимым структурам, Расположенным параллельно по отношению к сократительным элементам мышцы - Миофибриллам. К этим параллельным эластическим элементам относятся окружающие мышечное волокно оболочка (сарколемма), Элементы Соединительной ткани между волокнами и их пучками, и некоторые другие структурные компоненты мышечной клетки.
Зависимость между величиной внешней нагрузки и удлинением мышцы нелинейная. Модуль упругости покоящейся мышцы возрастает с ее растяжением.
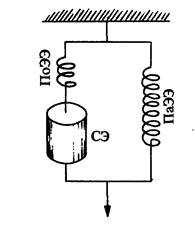
Рис. 6. Аналоговая модель строения мышцы: СЭ - сократительный элемент, ПаЭЭ - параллельный эластический элемент, ПоЭЭ - последовательный эластический элемент (по RueggJ. С. 1983).
Степень предварительного растяжения определяет и величину дополнительной силы Эластического напряжения мышцы, которую она может развить в процессе своей Активации при данной исходной длине. Соотношение сила - длина мышцы при ее Изометрическом Напряжении зависит от степени взаимного перекрывания нитей Актина и Миозина в Саркомере (рис.7). Максимум силы достигается при длине мышцы, соответствующей состоянию Саркомера, когда его длина составляет от 2,0 до 2,2 мкм. При меньшей длине мышцы (или Саркомера) сила уменьшается, поскольку Актиновые и Миозиновые нити начинают мешать друг другу, а также и из-за некоторых других факторов, возникающих при укорочении мышцы. Все это обычно не позволяет мышцам укорачиваться при сокращении до длины, меньшей, чем 50-70% от их длины в покое.
Если же мышцу растягивать более, чем ее Длина в покое, то сократительная сила также уменьшается, потому что нити Актина при этом вытягиваются из пучка нитей Миозина. Поэтому, при увеличении длины Саркомера до 2,9 мкм, мышечное волокно сможет развить только около 50% от своего Изометрического Максимума, так как зона взаимного перекрытия сократительных элементов составляет лишь половину от максимальной. При длине Саркомера 3,6 мкм и более Миофибриллы уже не могут генерировать силу, потому что их Актиновые и Миозиновые нити не перекрываются.
Мышцы на своих концах переходят в Сухожилия, через которые они передают усилия на Костные рычаги. Сухожилия также обладают упругими свойствами, которые классифицируются как Последовательный упругий элемент мышцы.
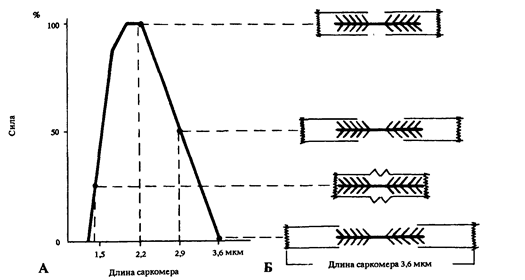
Рис. 7. Зависимость между силой сокращения, длиной саркомера и степенью перекрывания актиновых и миозиновых нитей: А - зависимость между максимальной изометрической силой, развиваемой саркомером во время тетануса и его длиной; Б - взаимное перекрывание актиновых и миозиновых нитей при различной длине саркомера (по Gordor A. M. и др., 1966, переработано).
При внезапном и сильном внешнем воздействии, или при высокой и резкой силе мышечного сокращения, Эластические элементы мышцы, растягиваясь, смягчают силовые воздействия, распределяя действие силы в течение более длительного промежутка времени. В некоторых случаях упругость мышц может создавать дополнительную силу, например, в начальной фазе движения.
Сухожилия обладают большей прочностью на растяжение (примерно 7000 Н/см2), чем мышечная ткань (всего около 60 Н/см2). Наиболее слабыми, и поэтому часто травмируемыми участками мышцы, являются Переходы мышцы в сухожилия. Поэтому, перед каждым тренировочным занятием, необходима хорошая предварительная подготовка разминка.
С биологической точки зрения физическая подготовка представляет собой процесс направленной адаптации организмах тренировочным воздействиям. Нагрузки, применяемые в процессе физической подготовки, выполняют, роль раздражителя, возбуждающего приспособительные изменения в организме. Тренировочный эффект определяется направленностью и величиной физиологических и биохимических изменений, происходящих под воздействием применяемых нагрузок. Глубина происходящих при этом в организме сдвигов зависит от основных характеристик физической нагрузки: Интенсивности и Продолжительности Выполняемых упражнений; Количества повторений упражнений; Вида физических упражнений; Продолжительности и Характера интервалов отдыха между повторением упражнений.
Определенное сочетание перечисленных параметров физических нагрузок приводит к необходимым изменениям в организме; к перестройке обмена веществ и, в конечном итоге, к росту тренированности.
Процесс адаптации организма к воздействию физических нагрузок имеет Фазный характер. Поэтому выделяют два этапа адаптации: срочный и долговременный (хронический).
Этап Срочной адаптации сводится преимущественно к изменениям энергетического обмена и связанных с ним функций вегетативного обеспечения на основе уже сформированных механизмов их реализации, и представляет собой непосредственный ответ организма на однократные воздействия физических нагрузок.
При многократном повторении физических воздействий и суммировании многих следов нагрузок, постепенно развивается Долгосрочная адаптация. Этот этап связан с формированием в организме функциональных и структурных изменений, происходящих вследствие стимуляции генетического аппарата нагружаемых во время работы клеток. В процессе долговременной адаптации к физическим нагрузкам активируется синтез нуклеиновых кислот и специфических белков, в результате чего происходит увеличение возможностей опорно-двигательного аппарата, совершенствуется его энергообеспечение.
Разовость протекания процессов адаптации к физическим нагрузкам позволяет выделять три разновидности эффектов в ответ на выполняемую работу.
Срочный тренировочный эффект, возникающий непосредственно во время выполнения физических упражнений и в период срочного восстановления в течение 0,5-1,0 часа после окончания работы. В это время происходит устранение образовавшегося во время работы кислородного долга.
Отставленный тренировочный эффект, сущность которого составляет активизация физической нагрузкой пластических процессов для избыточного синтеза разрушенных при работе клеточных структур и возобновление энергетических ресурсов организма. Этот эффект наблюдается на поздних фазах восстановления (обычно в пределах до 48 часов после окончания нагрузки).
Кумулятивный тренировочный эффект является результатом последовательного суммирования срочных и отставленных эффектов повторяющихся нагрузок. В результате кумуляции следовых процессов физических воздействий на протяжении длительных периодов тренировки (более одного месяца) происходит прирост показателей работоспособности и улучшение спортивных результатов.
Небольшие по объему физические нагрузки не стимулируют развитие тренируемой функции и считаются Неэффективными. Для достижения выраженного кумулятивного тренировочного эффекта необходимо выполнить объем работы, превышающий величину неэффективных нагрузок.
Дальнейшее наращивание объемов выполняемой работы сопровождается, до определенного предела, Пропорциональным увеличением тренируемой функции. Если же нагрузка превышает предельно допустимый уровень, то развивается Состояние перетренированности, происходит Срыв адаптации.
Похожие статьи
-
Наиболее подходящим возрастом для занятий атлетической гимнастикой является возраст от 16 до 18 лет, хотя нельзя отрицать и, занятия в более младшем...
-
У людей, которые систематически и активно занимаются физическими упражнениями, повышается психическая, умственная и эмоциональная устойчивость при...
-
ВЫВОДЫ - Основы периодизации круглогодичной тренировки
1. В результате анализа специальной научно-методической литературы по организационно - методическим основам построения тренировочных занятий атлетической...
-
При регулярных занятиях физическими упражнениями, каким-либо видом спорта увеличивается количество эритроцимышенной деятельности гемоглобина,...
-
Результаты исследования - Основы периодизации круглогодичной тренировки
Через три месяца, при подведении итогов, у занимающихся из экспериментальной группы было обнаружено, что объем грудной клетки у них в среднем, увеличился...
-
История развития ритмической гимнастики Ритмическая гимнастика начала развиваться еще в глубокой древности. Чтобы выжить в борьбе с природой,...
-
Содержание экспериментальной программы В связи с актуальностью вопроса об увеличении объема грудной клетки занимающихся атлетической гимнастикой и...
-
Организация исследования - Основы периодизации круглогодичной тренировки
Исследование проводилось на базе школьного тренажерного комплекса в период с 2003 по 2004 год и состояло из трех этапов. На первом этапе осуществлялось...
-
Задачи исследования Для достижения цели исследования необходимо было решить следующие задачи: 1. Исследовать по данным специальной научно-методической...
-
Физиологические функциональные системы (ФС) организма представляют собой динамические, саморегулирующиеся организации, все компоненты которых...
-
В циклических видах спорта может осуществляться любая мышечная деятельность, и в ней задействованы практически все группы мышц. Существует большое...
-
Формирование и совершенствование различных морфофизиологических функций и организма в целом зависят от их способности к дальнейшему развитию, что имеет...
-
Формирование и совершенствование различных морфофизиологических функций и организма в целом зависят от их способности к дальнейшему развитию, что имеет...
-
Одной из наиболее сложных проблем, возникающих при дополнительном введении в программу тренировок теннисиста упражнений по развитию силовых качеств,...
-
ВВЕДЕНИЕ - Основы периодизации круглогодичной тренировки
Актуальность . Атлетическая гимнастика - это вид спорта, способствующий укреплению здоровья, исправлению и лечению многих врожденных и приобретенных...
-
Форма и виды проявления силовых способностей Под силой подразумевают способность человека преодолевать внешнее сопротивление или противостоять ему за...
-
1. Фитнес - это образ жизни, который ведет к физическому и ментальному здоровью человека. Фитнес включает в свое понятие и занятия в тренажерном зале, и...
-
Мышечная работа (М. р.) - перемещение и поддержание положений тела и его частей благодаря работе мышц, обеспечиваемой координацией всех физиологических...
-
Обмен веществ - Биологические основы физической культуры
Сущность обмена веществ состоит в том, что из внешней среды в организм поступают богатые потенциальной энергией вещества, где они распадаются на более...
-
Физические упражнения выполняются с различной скоростью и величиной внешнего отягощения. Напряженность физиологических функций (интенсивность...
-
Дыхательная система - Биологические основы физической культуры
Дыхательная система включает воздухоносные пути, легкие, и другие органы, а также комплексы физиологических процессов, обеспечивающих потребление...
-
Тренировка - Бодибилдинг как спорт и образ жизни
- с помощью упражнений, направленных на всестороннее развитие всех мышечных групп, создание по возможности совершенного мышечного рельефа человеческого...
-
При систематических занятиях циклическими видами спорта улучшается кровоснабжение мозга, общее состояние нервной системы на всех ее уровнях. При этом...
-
Скелетная мускулатура - главный аппарат, при помощи которого совершаются физические упражнения. Хорошо развитая мускулатура является надежной опорой для...
-
При физической нагрузке потребление О2 и продукция СО2 возрастают в среднем в 15--20 раз. Одновременно усиливается вентиляция и ткани организма получают...
-
Биохимические закономерности адаптации к мышечной работе - Основы техники и тактики игры в волейбол
Любая физическая активность характеризуется своими специфическими двигательными действиями. Объединяет эти виды физической активности следующее: -...
-
Основы коррекции телосложения - Комплекс упражнений для женской гимнастики
Практически каждый человек стремиться выглядеть красиво и эстетично, быть здоровым и жить долго. Не последнее место занимает в достижении этих целей наше...
-
Общая характеристика выносливости Понятие "выносливость" издавна связывают со способностью человека продолжать более или менее эффективно совершать...
-
Задачи исследования 1. Изучить состояние проблемы о влиянии занятий атлетической гимнастикой на морфофункциональные показатели и силовые способности...
-
Под воздействием физических упражнений нормализуется состояние основных нервных процессов - повышается возбудимость при усилении процессов торможения,...
-
К управлению процессом самостоятельных занятий относится дозирование физической нагрузки, ее интенсивности на занятиях физическими упражнениями....
-
Заключение - Физическая подготовка спортсменов
Проблема изучения закономерностей физического развития человека давно привлекла к себе внимание исследователей. Так, например, еще в трудах П. Ф....
-
Основное средство физической культуры - физические упражнения. Существует физиологическая классификация упражнений, в которой вся многообразная мышечная...
-
Хотя точные механизмы этого явления пока не установлены, уже достаточно давно известно, что потребление углеводов во время двигательной активности...
-
Биологические принципы спортивной тренировки
Знание закономерностей развития адаптации к мышечной работе является обязательным условием грамотного, научно обоснованного построения тренировочного...
-
Использование различных средств физической культуры, занятия разными видами спорта тесно связаны с физиологическими закономерностями функционирования...
-
Сердечнососудистая система - Биологические основы физической культуры
Сердечнососудистая система (ССС) обеспечивает циркуляцию крови в организме и состоит из сердца и кровеносных сосудов. Кровь состоит на 55 % из жидкой...
-
Мышца -- это орган тела, состоящий из поперечно-полосатой или гладкой мышечной ткани, способной к сокращению под воздействием нервных импульсов....
-
Особенности тренировки самбистов - Организация тренировочного процесса самбистов
В последнее время борьба самбо приобрела особую популярность у молодежи. Практика работы лучших тренеров показывает, что из юных борцов выходит много...
-
Введение - Сила и основы методики ее воспитания
Под силой понимается способность человека преодолевать внешнее сопротивление или противодействовать ему за счет мышечных усилий. Один из наиболее...
ОБЗОР ЛИТЕРАТУРЫ ПО ТЕМЕ ИССЛЕДОВАНИЯ, Физиологические основы атлетической гимнастики - Основы периодизации круглогодичной тренировки