Конформационные изменения белков-гистонов - Эпигенетическая изменчивость
Как известно, молекула ДНК эукариот плотно упакована в микроскопическом объеме ядра, но при этом сохраняется возможность в нужный момент и в нужной комбинации извлекать из ДНК необходимую информацию, осуществлять эти противоположные задачи помогает хроматин.
Хроматин - чрезвычайно сложный и динамический комплекс, в состав которого входят ДНК и белки-гистоны, именно хроматин служит носителем генетической информации, участвую в избирательном и комбинаторном извлечении информации из ДНК. В зависимости от плотности упаковки различают: 1) гетерохроматин - плотно упакованный, хорошо виден под микроскопом, ДНК не транскрибируется. Встречается конститутивный (одинаков во всех типах клеток) и факультативный (отличается в разных типах клеток) гетерохроматин. 2) эухроматин - неплотно упакованный, слабо заметен под микроскопом, характеризуется транскрипционной активностью. Составляющей единицей хроматина является нуклеосома. Именно нуклеосома состоит из беков-гистонов и опоясывающей их молекулы ДНК.
Белки-гистоны - небольшие по молекулярной массе белки с высоким содержанием положительно заряженных аминокислотных остатков - лизинов и аргининов. Открыты А. Косселем в 1884 году. Молекула гистона состоит из центрального структурированного трехспирального домена и двух неструктурированных N - и С - хвостов. Существует пять типов гистонов: Н1, Н2А, Н2В, Н3 и Н4.
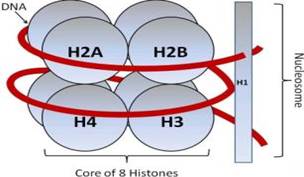
Рис. 4 - Строение нуклеосомы
Как видно на рисунке, нуклеосома состоит из восьми молекул гистонов: богатые лизином (Н2А)2 и (Н2В)2 и богатые аргинином (Н3)2 и (Н4)2, которые образуют октомер, вокруг которого навивается молекула ДНК. Н1-линкерный гистон, он связывается с межнуклеосомной ДНК.
В результате укладки гистонов, образуется 14 мотивов - белковых структур, расположенных на поверхности гистонов. Мотивы содержат АК+, которые обеспечивают контакт с отрицательно заряженными фосфатами остова ДНК. 14 бороздок ДНК втягивают в себя 14 мотивов, тем самым молекула ДНК жестко фиксируется на поверхности гистонов.
Хвосты гистонов выходят на поверхность хроматиновой фибриллы, они участвуют в межнуклеосомном взаимодействии, они очень подвижны и подвергаются многочисленным модификациям: ацетилирование (Лизин (К)),фосфолирирование (Треонин (Т), серин (S)), метилирование (Лизин (К), Аргинин (R)), убиквитинилирование (Лизин (К)), ADP-рибозилирование, сумоилирование.
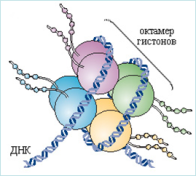
Рис. 5 - Хвосты гистонов, подвергающиеся модификациям
Ацетилирование гистоновых хвостов снижает положительный заряд гистонов, нейтрализует лизин(+), тем самым уменьшая притяжение между гистонами и ДНК. Ведет к активации транскрипции генов. Фосфолирирование гистонов совпадает с началом митоза, который начинается с фосфорилирования линкерного гистона H1. Хвосты (N-концевые части) коровых гистонов подвергаются фосфорилированию во время деления клетки. Метилирование - не изменяет заряд гистонов, существует моно-,ди - и триметилирование. Модификации хвостов гистоновых белков влияют на упаковку хроматина, на его поведение при активации генов, их репрессии и многих других процессов, связанных с доступом к ДНК. Наиболее разработанной моделью функционирования хроматина считается "гистоновый код".
"Гистоновый код" - разнообразный набор модификаций гистоновых хвостов, который можно целенаправлено менять и передавать по наследству. Он и определяет функциональное состояние гена. Является основным эпигенетическим механизмом, контролирующим включение или выключение генов и передачу этой программы контроля по наследству от клетки к клетке.
Похожие статьи
-
Выделяют два основных уровня контроля активации и ингибирования генов - химическую модификацию ДНК и конформационные изменения белков-гистонов....
-
Этапы биосинтеза белка, Транскрипция - Биосинтез белков. Ген и его роль в синтезе белков
Транскрипция Мост между геном (кодонами) и белком обеспечивается РНК. Точнее, информация, закодированная в последовательности азотистых оснований ДНК,...
-
Введение - Биосинтез белков. Ген и его роль в синтезе белков
Способность клеток поддерживать высокую упорядоченность своей организации в хаотичной Вселенной зависит от генетической информации, которая реализуется,...
-
Введение, Эпигенетика. Что такое эпигенетическая изменчивость - Эпигенетическая изменчивость
В геноме человека содержится около 25 тыс. генов, но одновременно в клетке любой живой ткани работает не более половины всех генов. Часть из них нужна...
-
Заключение - Биосинтез белков. Ген и его роль в синтезе белков
В ходе написания курсовой работы мне удалось выяснить что биосинтез белков осуществляется во всех клетках про - и эукариот. Информация о первичной...
-
В цитозоле клеток 20 различных аминокислот присоединяются б-карбоксильной группой к 3'-гидроксильному акцепторному концу соответствующих тРНК с...
-
Регуляция синтеза гема, Нарушения синтеза гема - Порфирии - Первичная структура белков
Регуляторными ферментами являются АЛК (аминолевулинатсинтетаза) и Аминолевулинатдегидрогеназа. АЛК - синтаза является главным регуляторным ферментом,...
-
Замена аминокислоты на поверхности гемоглобина А - Первичная структура белков
В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 в-цепи, в которых глутамат, высокополярная отрицательно заряженная...
-
Трансляция - Биосинтез белков. Ген и его роль в синтезе белков
Трансляция - важная составная часть общего метаболизма, и ее сущность заключается в переводе генетической информации с мРНК, являющейся первичным...
-
Изменения свойств поверхности - Строгие постулаты Коха
Заражение клеток трансформирующими вирусами приводит не только к упомянутым выше изменениям их морфологии и способности к размножению, но и к резко...
-
Обмен белков - Роль белков в организме
Аминокислоты, образовавшиеся после расщепления белков в пищеварительном тракте, всасываются в кровь. В кровь всасывается также незначительное количество...
-
Обмен белков в организме человека - Строение, функции и значение белков
Важный критерий пищевой ценности белков - доступность аминокислот. Аминокислоты большинства животных белков полностью высвобождаются в процессе...
-
Обмен белков - Питательные вещества
После расщепления белков в пищеварительном тракте образовавшиеся аминокислоты всасываются в кровь. В кровь всасывается также незначительное количество...
-
Свойство белков - Основы генетики
Белки - высокомолекулярные соединения, построенные из аминокислот и являются одними из наиболее сложных по строению и составу среди всех органических...
-
Азотистое равновесие - Строение, функции и значение белков
Азотистое равновесие - это соотношение между количеством азота, содержащегося в принятой пище, и количеством азота, выведенного из организма. Если обе...
-
Роль белков в организме - Роль белков в организме
Функции белков в организме разнообразны. Они в значительной мере обусловлены сложностью и разнообразием форм и состава самих белков. Белки -- незаменимый...
-
Классификация белков - Роль белков в организме
Белки подразделяются на две большие группы: простые белки, или протеины, и сложные белки, или протеиды. При гидролизе протеинов в кислом водном растворе...
-
Строение, свойства и функции - Роль белков в организме
"Во всех растениях и животных присутствует некое вещество, которое, без сомнения, является наиболее важным из всех известных веществ живой природы, и без...
-
Введение - Роль белков в организме
Нормальная деятельность организма возможна при непрерывном поступлении пищи. Входящие в состав пищи жиры, белки, углеводы, минеральные соли, вода и...
-
Ген и его строение - Биосинтез белков. Ген и его роль в синтезе белков
Ген представляет собой элементарную единицу функции наследственного материала. Это означает, что фрагмент молекулы ДНК, соответствующий отдельному гену и...
-
Физиологическая роль белка, Структурная функция белка - Строение, функции и значение белков
Структурная функция белка Белки сложные органические соединения, построенные из аминокислот. В состав белковых молекул входят азот, углерод, водород и...
-
Введение - Строение, функции и значение белков
Важнейшим компонентом питания являются белки. Белки представляют основу структурных элементов клетки и тканей. С белками связаны основные проявления...
-
Изменения объема легких - Функции дыхательной системы
У человека легкие занимают около 6% объема тела независимо от его веса. Объем легкого меняется при вдохе не всюду одинаково. Для этого имеются три...
-
Кариоплазма, Хроматин - Клеточное ядро
Кариоплазма (ядерный сок, нуклеоплазма) - основная внутренняя среда ядра, она занимает все пространство между ядрышком, хроматином, мембранами,...
-
Развитие половой системы - длительный процесс, который не завершается в эмбриональном периоде, а продолжается после рождения, вплоть до достижения...
-
Для синтеза полипептидной цепи необходимо большое количество компонентов, совместное и согласованное взаимодействие которых приводит к образованию белка....
-
Классификация белков - Питательные вещества
Белки подразделяются на две большие группы: простые белки, или протеины, и сложные белки, или протеиды. При гидролизе протеинов в кислом водном растворе...
-
Функции белков в организме - Питательные вещества
Функции белков в организме разнообразны. Они в значительной мере обусловлены сложностью и разнообразием форм и состава самих белков. Белки - незаменимый...
-
Ген. Взаимосвязь между геном и признаком - Биосинтез белков. Ген и его роль в синтезе белков
Долгое время ген рассматривали как минимальную часть наследственного материала (генома), обеспечивающую развитие определенного признака у организмов...
-
Вода и растворенные в ней вещества, числе минеральные соли, создают внутреннюю среду организма, свойства к-й сохраняются постоянными или изменяются...
-
Значение белков в питании - Белок в жизни живых организмов
Белок - необходимая составная часть продуктов питания. Проблема пищевого белка стоит очень остро. По данным Международной организации по продовольствию и...
-
- создают онкотическое давление крови, от которого зависит обмен воды между кровью и межклеточной жидкостью; - определяют вязкость крови, что в свою...
-
Люди впервые получили инсулин в 1921 году из поджелудочной железы собак Бантинг и Бест приготовили экстракт. В кристаллическом виде в 1926 году Sanger...
-
Патологическая физиология - Изменение при сахарном диабете
Поджелудочная железа как основной источник заболевания В 1869 году Пауль Лангерганс гистологически идентифицировал островковые клетки, составляющие...
-
Характеристика модификационной изменчивости - Формы изменчивости живых организмов
-обратимость -- изменения исчезают при смене специфических условий окружающей среды, спровоцировавших их -групповой характер -изменения в фенотипе не...
-
Первичная структура - понятие, обозначающее последовательность аминокислотных остатков в белке Пептидная связь - основной вид связи, определяющий...
-
Аутоиммунные процессы, Ожирение и питание - Изменение при сахарном диабете
На возможность значения аутоиммунного процесса (Nerup J. et all, 1978) в развитии диабета указывают ряд данных: 1) присутствие в островках поджелудочной...
-
Непременным компонентом вирусной частицы является какая-либо одна из двух нуклеиновых кислот, белок и зольные элементы. Эти три компонента являются...
-
Роль разных форм изменчивости в эволюции видов - Чарльз Дарвин о формах и причинах изменчивости
В то время, когда работал Дарвин, явления изменчивости организмов были изучены очень слабо, однако ему было ясно, что эволюционный процесс базируется на...
-
Нарушение аминокислотного обмена - Строение, функции и значение белков
Наиболее часто встречающимися болезнями, связанными с нарушением аминокислотного обмена, являются фенилкетонурия и альбинизм. В норме аминокислота...
Конформационные изменения белков-гистонов - Эпигенетическая изменчивость