Терминация - Молекулярный механизм репликации ДНК
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются недореплицированными. Это происходит потому, что ДНК-полимераза в, отвечающая за заполнение бреши, образованной после удаления праймера, не может вести синтез цепи ДНК от 3'- к 5'-концу (рис. А).
Таким образом, в ходе каждого цикла репликации 5'-концы синтезированных цепей укорачиваются. Но такие потери не представляют опасности для генетической информации хромосом, потому что укорочение ДНК идет за счет теломер. Во время следующего цикла репликации 5'-концы цепей ДНК опять остаются недостроенными. Таким образом, с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочение теломер в большинстве клеток по мере их старения - важный фактор, определяющий продолжительность жизни организма.
Однако в эмбриональных и других быстроделящихся клетках потери концов хромосом недопустимы, потому что укорочение ДНК будет происходить очень быстро. В эукариотических клетках имеется фермент теломераза (нуклеотидилтрансфераза), обеспечивающий восстановление недореплицированных 5'-концов. К особенностям этого фермента относят присутствие в качестве простетической группы РНК. Фрагмент РНК в активном центре теломеразы служит матрицей при синтезе теломер-ных повторов хромосом.
С помощью РНК фермент комплементарно прикрепляется к 3'-концу недостроенной дочерней цепи ДНК. Теломераза по принципу комплементарности последовательно удлиняет 3'-конец цепи ДНК на один гексануклеотид - GGGTTA-. Синтез всегда идет от 5'- к 3'-концу. Затем теломераза смещается по цепи ДНК на один теломер и начинает синтез нового фрагмента - GGGTTA - (рис. Б).
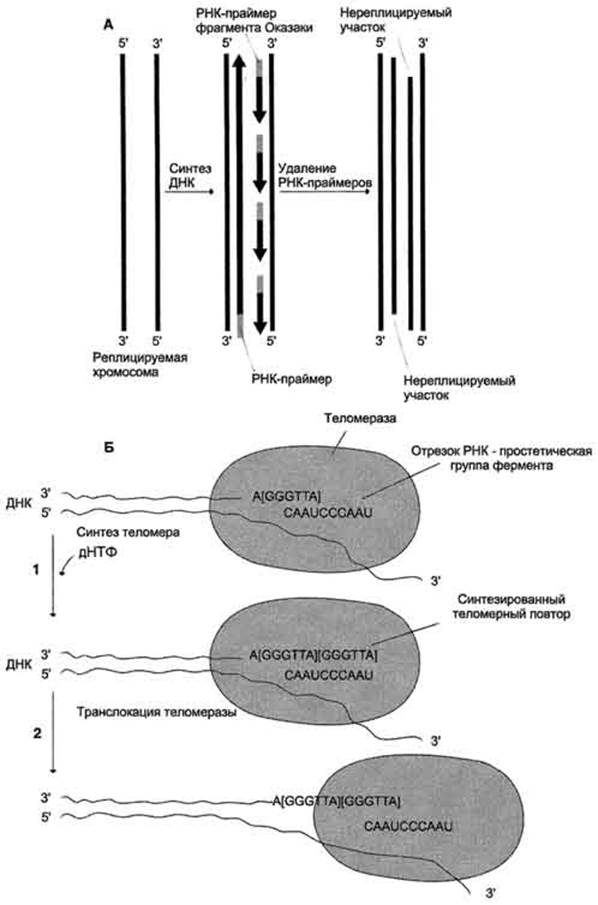
А - на рисунке показано укорочение вновь синтезированных цепей ДНК после удаления праймеров; Б - в состав теломеразы входит короткая молекула РНК, содержащая в активном центре последовательность нук-леотидов, комплементарную теломерному повтору; 1 - фермент прикрепляется за счет взаимодействия РНК с существующей теломерой и добавляет последовательно по одному нуклеотиду фрагмент - GGGTTA-. Матрицей служит простетическая группа теломеразы - фрагмент РНК; 2 - фермент перемещается по нити ДНК таким образом, что РНК-матрица в составе теломеразы постоянно комплементарно связана с концом вновь синтезированного теломерного повтора. Заново синтезированная тело-мерная ДНК служит матрицей для удлинения второй цепи ДНК, но уже в ходе следующего цикла клеточного деления. Теломер-ный повтор на рисунке взят в квадратные скобки -[GGGTTA]-.
В большинстве соматических клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и ее потомства. Однако небольшую активность теломеразы обнаруживают в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия, эпидермиса кожи и др.
Похожие статьи
-
Элонгация - Молекулярный механизм репликации ДНК
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами. Данный фермент обладает некоторыми свойствами: 1. Фермент не может начинать синтез. Он не...
-
Инициация репликации - Молекулярный механизм репликации ДНК
Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы - факторы роста....
-
К чистым факторам переноса относят, например, факторы F и F-подобные (pAP22-4, pAP38, pAP39, pAP41) и pTRA1, фактор T, идентифицированный в E. coli,...
-
Молекулярные механизмы регуляции клеточной смерти - Биологические механизмы жизненного цикла клетки
Апоптоз - многоэтапный процесс. Первый этап - прием сигнала, предвестника гибели в виде информации, поступающей к клетке извне или возникающей в недрах...
-
Молекулярный механизм сокращения - Биофизика мышечного сокращения
Один грамм ткани скелетной мышцы содержит примерно 100 мг "сократительных белков" - актина и миозина. Механизм их взаимодействия во время элементарного...
-
Молекулярная основа устойчивости клеток к вирусам. - Строгие постулаты Коха
Хотя устойчивость, индуциорованная интерфероном, защищает клетки от самых различных ДНК РНК-вирусов, степень защиты от разных вирусов неодинакова. Кроме...
-
Некротическая, беспрограммная форма гибели наиболее характерна для сильных воздействий, например, для облучения клеток и организма в высоких, летальных...
-
Как управлять экспрессией - Технологии изучения клеточных механизмов памяти
Понимание того, каким же образом сигнал, поступающий к клетке, передается на немедленные ранние гены и приводит к их активации, позволяет влиять на их...
-
Жизненный цикл клетки - Биологические механизмы жизненного цикла клетки
Биологический клетка деление До сих пор много тайн клетки остаются неразгаданными. Загадочным во многом остается и запрограммированный генетически...
-
Молекулярная и генетическая организация плазмид - Молекулярная и генетическая организация плазмид
Генетическая организация разных плазмид отличается большим разнообразием, так как среди плазмид, на основе их функциональной специфичности, различают...
-
Классификация плазмид, Распространение плазмид - Молекулярная и генетическая организация плазмид
В 50-е гг. плазмиды R стали классифицировать на fi+ и fi - (по способности ингибировать перенос плазмиды F). Далее стали, в зависимости от различия в...
-
История исследования плазмид - Молекулярная и генетическая организация плазмид
Начало исследования плазмид относят к 20 гг. XX века. В 1921 г. Bourdet и Ciuca открыли лизогенные бактерии, способные спонтанно лизироваться. В 1925 г....
-
Введение - Молекулярная и генетическая организация плазмид
Плазмиды - внехромосомные генетические элементы, способные к автономному поддержанию в цитоплазме бактерий или существованию в нтегрированном в хромосому...
-
Механизм экзоцитоза медиатора - Нервно-мышечная физиология
Для экзоцитоза необходимы ионы кальция, которые входят в терминаль через кальциевые каналы и белки экзоцитоза. Среди них синаптотагмин, синаптобревин,...
-
Механизм действия гормонов. Роль циклазной системы в механизме действия гормонов - Понятие гормонов
По механизму действия гормоны делят на два основные типа. Первый - это белковые и пептидные гормоны, катехоламины и гормоноиды. Их молекула, подойдя к...
-
Биологические виды, Изоляция и видообразование - Механизмы эволюции
Вплоть до XVII в. исследователи опирались на представление о виде, созданное еще Аристотелем, который воспринимал виды как совокупности сходных особей....
-
Органы-мишени и механизм действия инсулина - Биохимия поджелудочной железы
1. В крови инсулин может быть в двух формах: свободной и связной. Органы - мишени свободного инсулина: печень, скелетная мускулатура, кишечник; связного...
-
Механизмы апоптоза - Радиационный апоптоз и некроз клеток облученного организма
Развитие и функционирование многоклеточных организмов подразумевает равновесие между синтезом и распадом молекул, их окислением и восстановлением,...
-
Как увидеть память - Технологии изучения клеточных механизмов памяти
Итак, экспрессия многих генов зависит от нейронной активности, соответственно, этот феномен получил название активностно-зависимой генной экспрессии. А...
-
Бактериоцины - вещества, летальные для клеток бактерий. Их названия определяются названиями микроорганизмов-продуцентов. Это термостабильные белки,...
-
Чем занимаются немедленные ранние гены в клетке? - Технологии изучения клеточных механизмов памяти
Некоторые из них кодируют транскрипционные факторы для структурных генов, другими словами, регулируют уровни их экспрессии [3]. Это очень важная функция,...
-
Молекулярный состав клетки, Неорганические вещества. - Химический состав клетки
Химический элементы входят в состав клеток в виде ионов или компонентов молекул неорганических и органических веществ. Неорганические вещества. Вода -...
-
Механизмы доступа к БД - Телемедицина
Существует, в общем, два класса механизма доступа к БД: - на стороне сервера (используются интерфейсы CGI, API и др.); - на стороне клиента (используются...
-
Несовместимость и группы несовместимости - Молекулярная и генетическая организация плазмид
К одной группе несовместимости (inc-группа) относят плазмиды, которые несовместимы между собой, но совместимы с любой плазмидой из другой группы....
-
Заключение - Биосинтез белков. Ген и его роль в синтезе белков
В ходе написания курсовой работы мне удалось выяснить что биосинтез белков осуществляется во всех клетках про - и эукариот. Информация о первичной...
-
Биосинтез РНК(Транскрипция) - Регуляция экспрессии генов у прокариот
Синтез РНК можно представить следующей схемой : Субстратами реакции служат трифосфаты рибонуклеозидов. Реакция идет только в присутствии ДНК, выполняющей...
-
Механизм дыхательных движений - Строение и функции дыхательной системы
Газообмен в легких происходит в результате ритмических дыхательных движений-- вдоха и выдоха. У взрослого человека происходит 16-18 дыхательных движений...
-
Механизм расслабления - Нервно-мышечная физиология
Начинается с уменьшения концентрации ионов кальция в саркоплазме, за счет работы кальциевых насосов и натрий-кальциевого обменника, разрушение...
-
В процессе хемомеханического преобразования АТФ обеспечивает разъединение головки миозина и актинового филамента и энергетику для дальнейшего...
-
Блокирование нмс 1 уровень - пресинаптический - а) можно заблокировать ПД по мембране, блокаторами натриевых, калиевых каналов: тетродотоксин,...
-
Механизмы и этапы формирования биопленок Биопленка - микробное сообщество, характеризующееся клетками, которые прикреплены к поверхности или друг к...
-
Механизмы морфогенетических движений в процессе гаструляции. - Гаструляция и нейруляция
Несмотря на все разнообразие описанных выше гаструляционных морфогенетических движений, все они, ровно, как и морфогенетические движения, в последующем...
-
Как управлять памятью - Технологии изучения клеточных механизмов памяти
Что, если создать трансгенных мышей, у которых, как и у уже рассмотренных, промотор немедленных ранних генов находится под контролем системы tTA-tetO или...
-
Врожденные защитные механизмы - Особенности нервной системы у детей
Новорожденные далеко не так беззащитны, как это может показаться со стороны. Природа наделила детей первых месяцев жизни мощными врожденными механизмами,...
-
Масляно-кислое брожение -- это сложный биохимический процесс превращения сахара масляно-кислыми бактериями в анаэробных условиях с образованием масляной...
-
Вместе с тем в их организации есть и отличия, которые обусловлены характером результата. ФС, определяющие различные показатели внутренней среды...
-
Плоскостопие - Своды стопы, механизмы укрепляющие своды стопы. Плоскостопие
Виды плоскостопия В соответствии с причинами, из-за которых происходит уплощение стопы, плоскостопие делится на пять основных видов. У большинства...
-
Биохимические особенности стареющего организма, Основные механизмы старения - Возрастная биохимия
Основные механизмы старения Биология старения - раздел геронтологии, который изучает старение живых организмов на всех уровнях их организации....
-
Физиологические механизмы памяти - Биосоциальная сущность человека
Для адаптации индивидуального поведения к окружающей среде, тем более для осуществления разнообразной высшей нервной деятельности, человеку, равно как и...
-
Виды клеточной смерти - Биологические механизмы жизненного цикла клетки
Гибель (смерть) отдельных клеток или целых их групп постоянно встречается у многоклеточных организмов, также как гибель одноклеточных организмов. Причины...
Терминация - Молекулярный механизм репликации ДНК