Молекулярный механизм сокращения - Биофизика мышечного сокращения
Один грамм ткани скелетной мышцы содержит примерно 100 мг "сократительных белков" - актина и миозина.
Механизм их взаимодействия во время элементарного акта мышечного сокращения объясняет теория скользящих нитей, разработанная Хаксли и Хансон.
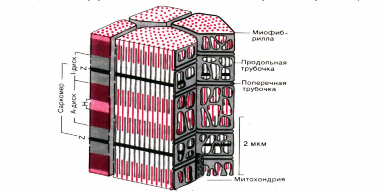
Рис. 1. Схема участка волокна скелетной мышцы человека (по Garamvolgyi)
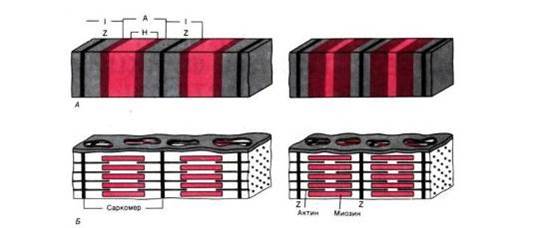
Рис. 2. А. оперечнополосатая структура миофибрилл: слева расслабление, справа сокращение. Б. Организация миозиновых и актиновых нитей в расслабленном и сократившемся саркомере. Аддитивный характер укорочения последовательно соединенных саркомеров
Теория скользящих нитей.
Сократительные белки актин и миозин образуют в миофибриллах тонкие и толстые миофиламенты. Они располагаются параллельно друг другу внутри мышечной клетки. Миофибриллы представляют собой сократимые пучки "нитей" (филаментов) диаметром около 1 мкм. Перегородки, называемые Z-пластинками, разделяют их на несколько компартментов-саркомеров длиной примерно по 2,5 мкм.
Согласно теории Хаксли и Хансон поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч "толстых" нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 "тонких" (толщиной по 5 нм) нитей актина, прикрепленных к Z-пластинкам наподобие щетинок в щетке. Пучок лежащих в определенном порядке миозиновых нитей длиной 1,6 мкм в середине саркомера выглядит в световом микроскопе темной полосой шириной 1,6 мкм; из-за свойства двойного лучепреломления в поляризованном свете (т. е. анизотропии) она называется А-диском. По обе стороны от А-диска находятся изотропные участки, содержащие только тонкие нити и поэтому выглядящие светлыми; эти так называемые I-диски тянутся до Z-пластинок. Именно в результате такого периодического чередования светлых и темных полос в саркомерах миофибриллы сердечной и скелетной мускулатуры выглядят поперечно-полосатыми.
В покоящейся мышце концы толстых и тонких филаментов обычно лишь слабо перекрываются на границе между А - и I-дисками. Эта зона перекрывания в А-диске выглядит в световом микроскопе гораздо темнее центральной Н-зоны, в которой нет актиновых нитей. На электронных микрофотографиях Н-зоны видна очень тонкая темная М-линия в середине саркомера - сеть опорных белков, по-видимому, удерживающих толстые нити в составе единого пучка.
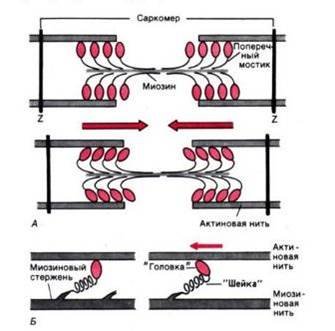
Рис. 3. Функция поперечных мостиков. А. Модель механизма сокращения: миозиновая нить с поперечными мостиками, прикрепленными к соседним актиновым нитям; вверху-до, внизу-после "гребка" мостиков (на самом деле они функционируют асинхронно). Б. Модель механизма генерирования силы поперечными мостиками; слева до, справа - после "гребка". Поперечные мостики химически соответствуют субфрагменту миозина - тяжелому меромиозину", который состоит из субфрагментов I (головка) и II (шейка)
Укорочение саркомеров. Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах. Сравнивая структуры саркомера в двух различных функциональных состояниях, можно видеть изменения поперечной исчерченности и взаиморасположения нитей во время сокращения: тонкие актиновые филаменты скользят вдоль толстых миозиновых, двигаясь между ними к середине их пучка и саркомера.
Длина нитей не меняется и при растяжении мышцы. Тонкие филаменты попросту вытягиваются из промежутков между толстыми нитями, так что степень перекрывания их пучков уменьшается.
Работа поперечных мостиков. Миозиновые нити несут поперечные, отходящие биполярно, как показано на рис. 3, А, выступы длиной около 20 нм с головками примерно из 150 молекул миозина. Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми (рис. 3, А). Движение головок создает объединенное усилие, как бы "гребок", продвигающий актиновые нити к середине саркомера. Сама биполярная организация молекул миозина обеспечивает противоположную направленность (стрелки на рис. 3) скольжения актиновых нитей в левой и правой половинах саркомера.
В результате однократного движения поперечных мостиков вдоль актиновой нити саркомер укорачивается только на 2 х 10 нм, т. е. примерно на 1% своей длины. Однако при изотоническом сокращении мышцы лягушки саркомеры за десятую долю секунды укорачиваются на 0,4 мкм, т. е. на 20% длины. Для этого поперечные мостики должны совершить свои гребковые движения за указанный промежуток времени не один, а 20 раз.
Генерирование мышечной силы. Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга, т. е. в строго изометрических экспериментальных условиях. Рис. 3, Б иллюстрирует такой процесс генерирования изометрической силы. Сначала головка миозиновой молекулы (поперечный мостик) прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом примерно 45°, возможно, благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика (видимо, "шейки" между головкой и миозиновой нитью) в напряженное состояние. Возникающее в результате упругое растяжение достигает лишь около 10 нм. Упругое натяжение, создаваемое индивидуальным поперечным мостиком, так слабо, что для развития мышечной силы, равной 1 мН, нужно объединить усилия по крайней мере миллиарда таких соединенных параллельно мостиков.
Даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряженном состоянии. На самом деле каждая миозиновая головка уже через сотые или десятые доли секунды отделяется от актиновой нити; однако через такое же короткое время следует новое прикрепление к ней. Несмотря на ритмичное чередование прикреплений и отделений с частотой порядка 5-50 Гц, сила, развиваемая мышцей в физиологических условиях, остается неизменной (исключение-летательные мышцы насекомых), так как статистически в каждый момент времени в прикрепленном, обусловливающем напряжение, состоянии находится одно и то же количество мостиков.
Изометрическое теплообразование. Мышца, поддерживающая определенное сократительное напряжение в изометрических условиях, отличается от сокращающейся изотонически тем, что не выполняет внешней работы (произведение силы на расстояние равно нулю). Однако в каждом цикле прикрепления-отделения поперечных мостиков совершается внутренняя работа по растяжению их упругих структур, которая преобразуется в тепло в момент отсоединения миозиновых головок. Теплота изометрического сокращения ("изометрическая работа") за единицу времени возрастает с увеличением количества функционирующих поперечных мостиков и частоты "гребков", требующих расхода АТФ.
Похожие статьи
-
Блокирование нмс 1 уровень - пресинаптический - а) можно заблокировать ПД по мембране, блокаторами натриевых, калиевых каналов: тетродотоксин,...
-
Введение - Биофизика мышечного сокращения
Физика и биология, на первый взгляд, довольно далекие друг от друга науки. Но это только на первый взгляд. В действительности же в этих науках есть много...
-
Регуляция мышечного сокращения - Биофизика мышечного сокращения
Обычно мышца возбуждается при поступлении потенциалов действия от иннервирующих мотонейронов; в результате передачи возбуждения через нервно-мышечные...
-
Присоединение Са 2+ к тропонину устраняет постоянно существующее препятствие для взаимодействия поперечных мостиков с актином. Из результатов...
-
В гладкой мышце присутствуют 2 типа сопряжения между процессами возбуждения и сокращения - электромеханический тип и фармомеханисеский тип, возникающий...
-
В естественных условиях одиночные сокращения скелетных мышц не встречаются. Происходит сложение, или суперпозиция, сокращений отдельных нейромоторных...
-
В процессе хемомеханического преобразования АТФ обеспечивает разъединение головки миозина и актинового филамента и энергетику для дальнейшего...
-
Строение скелетных мышц Каждая мышца состоит из параллельных пучков поперечнополосатых мышечных волокон. Каждый пучок одет оболочкой. И вся мышца снаружи...
-
1) [-]продолговатый мозг 2) [-]промежуточный мозг 3) [+]большие полушария переднего мозга, мозжечок 4) [-]средний мозг Из какого белка состоит оперенье...
-
Терминация - Молекулярный механизм репликации ДНК
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются...
-
Гладкие мышцы - Заболевания и травмы скелета человека. Заболевания мышечной системы человека
Кроме скелетных мышц, в нашем организме в соединительной ткани находятся гладкие мышцы в виде одиночных клеток. В отдельных местах они собраны в пучки....
-
К чистым факторам переноса относят, например, факторы F и F-подобные (pAP22-4, pAP38, pAP39, pAP41) и pTRA1, фактор T, идентифицированный в E. coli,...
-
Элонгация - Молекулярный механизм репликации ДНК
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами. Данный фермент обладает некоторыми свойствами: 1. Фермент не может начинать синтез. Он не...
-
Инициация репликации - Молекулярный механизм репликации ДНК
Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы - факторы роста....
-
Молекулярные механизмы регуляции клеточной смерти - Биологические механизмы жизненного цикла клетки
Апоптоз - многоэтапный процесс. Первый этап - прием сигнала, предвестника гибели в виде информации, поступающей к клетке извне или возникающей в недрах...
-
Мышечная ткань - Анатомия, конспект лекций
Охарактеризовать мышечную ткань, как ткань, обладающую способностью к сокращению, что позволяет осуществлять двигательные процессы в организме. Отметить...
-
Одиночное мышечное сокращение возникает в ответ на одиночную стимуляцию Различают периоды 1 латентный период - с момента начала стимуляции до момента...
-
Работа мышц - Заболевания и травмы скелета человека. Заболевания мышечной системы человека
Мышцы, сокращаясь или напрягаясь, производят работу. Она может выражаться в перемещении тела или его частей. Такая работа совершается при поднятии...
-
Стопа устроена и функционирует как упругий подвижный свод. Сводчатое строение стопы отсутствует у всех животных, включая антропоидов, и является...
-
Строение внутренней оболочки сердца эндокарда - Характеристика сердечной мышечной ткани
Эндокард выстилает изнутри камеры сердца, папиллярные мышцы, сухожильные нити и клапаны сердца. Толщина эндокарда в различных участках неодинакова: толще...
-
Механизм расслабления - Нервно-мышечная физиология
Начинается с уменьшения концентрации ионов кальция в саркоплазме, за счет работы кальциевых насосов и натрий-кальциевого обменника, разрушение...
-
Адаптационные изменения в сердечно-сосудистой системе. Сердце, адаптированное к физической нагрузке, обладает высокой сократительной способностью. Но оно...
-
Механизм дыхательных движений - Строение и функции дыхательной системы
Газообмен в легких происходит в результате ритмических дыхательных движений-- вдоха и выдоха. У взрослого человека происходит 16-18 дыхательных движений...
-
Сокращаясь, мышцы выполняют работу. Работа зависит от их силы. Мышца тем сильнее, чем больше в ней мышечных волокон, то есть чем она толще. При пересчете...
-
Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием белка миоглобина, который способен связывать...
-
Формирование скелетных мышц происходит на очень ранних этапах развития. На восьмой неделе внутриутробного развития различимы уже все мышцы, а к десятой...
-
- миографии - реографии - определения диаметра мышц и нервов + определения порога раздражения - динамометрии К физиологическим свойствам возбудимых...
-
Соединение костей, Мышечная система - Анатомия, конспект лекций
Объяснить, что в скелете человека в результате сочленения костей образуются либо механически более прочные непрерывные соединения, либо подвижные...
-
Патологическая гистология сердечной мышечной ткани - Характеристика сердечной мышечной ткани
Различные повреждающие воздействия на сердце (прекращение притока артериальной крови, травмы, воспаления и др.) могут вызвать некроз мышечной ткани, т....
-
Направления эволюции, Генетические и онтогенетические основы эволюции - Механизмы эволюции
Направление эволюции каждой систематической группы определяется взаимоотношениями между особенностями среды, в которой протекает эволюция данного...
-
Введение, Общая характеристика сердечной мышечной ткани - Характеристика сердечной мышечной ткани
В современной медицине все больший интерес вызывают вопросы лечения и профилактики заболеваний сердечно-сосудистой системы, возникновение которых в...
-
Биологические виды, Изоляция и видообразование - Механизмы эволюции
Вплоть до XVII в. исследователи опирались на представление о виде, созданное еще Аристотелем, который воспринимал виды как совокупности сходных особей....
-
Мышечная дистрофия - одно из самых тяжелых заболеваний опронодвигательного аппарата. В настоящее время нет лекарства на основе стволовых клеток для...
-
В сфере научных интересов сразу несколько направлений. Во-первых, применение стволовых клеток пуповинной крови и костного мозга при нейродегенеративных...
-
1) [-]земноводные 2) [-]рыбы 3) [+]ланцетник 4) [-]пресмыкающиеся Укажите правильную последовательность строения передней конечности лягушки: 1) ключица;...
-
Функциональная система адаптации - Механизмы адаптации организма человека к двигательной нагрузке
Проведенные в последние годы исследования механизмов и закономерностей адаптации людей к различным условиям деятельности привелидоказывают, что...
-
Механизм наркотической толерантности - Алкоголь
Многие психоактивные наркотики химически подобные нейротрансмиттерам - веществам, выделяемым нервными окончаниями при стимулирующем импульсе....
-
Введение - Заболевания и травмы скелета человека. Заболевания мышечной системы человека
Костно-мышечная система формируется в организме человека одной из первых. Именно она становится тем каркасом, на котором, словно на оси детской...
-
К двигательным качествам относятся сила, скорость (быстрота), ловкость, гибкость и выносливость. Они обусловлены ростом и развитием всего организма....
-
Плоскостопие - Своды стопы, механизмы укрепляющие своды стопы. Плоскостопие
Виды плоскостопия В соответствии с причинами, из-за которых происходит уплощение стопы, плоскостопие делится на пять основных видов. У большинства...
Молекулярный механизм сокращения - Биофизика мышечного сокращения