Регуляция мышечного сокращения - Биофизика мышечного сокращения
Обычно мышца возбуждается при поступлении потенциалов действия от иннервирующих мотонейронов; в результате передачи возбуждения через нервно-мышечные синапсы генерируются мышечные потенциалы действия (непрямая стимуляция). Возможна и прямая стимуляция мышечных волокон, но только в экспериментальных условиях. Например, при раздражении изолированной мышцы лягушки одиночным электрическим импульсом длительностью около 1 мс по мышечному волокну от места раздражения примерно через 1-2 мс со скоростью примерно 2 м/с будет распространяться потенциал действия, а еще через несколько миллисекунд оно сократится. Таким образом, сокращение вызывается потенциалом действия, т. е. возбуждением мембраны волокна.
Электромеханическое сопряжение.
Передача команды к сокращению от возбужденной клеточной мембраны к миофибриллам в глубине клетки (электромеханическое сопряжение) включает в себя несколько последовательных процессов, ключевую роль в которых играют ионы Са2+.
Локализация и механизм действия Са2+. Инъекция Са2+ в мышечные волокна вызывает их сокращение. Интактные живые волокна гораздо меньше подходят для демонстрации прямого воздействия Са2+ на миофибриллы, чем те же волокна после удаления или разрушения поверхностной клеточной мембраны. Для этого их либо "обдирают" ("скинируют") механически, либо обрабатывают детергентами, либо используют упоминавшееся выше экстрагирование глицеролом. Такие лишенные сарколеммы ("скинированные") мышечные волокна сокращаются только при погружении в раствор, содержащий АТФ и по крайней мере 10-6М ионизированного кальция для активации АТФазы. В этих условиях поперечные мостики миозиновых нитей могут за счет постоянного расщепления АТФ циклически взаимодействовать с актиновыми нитями. Если активирующий фактор Са2+ удалить из среды (например, добавив связывающие его вещества), миофибриллы расслабляются, поскольку взаимодействие между поперечными мостиками и актином предотвращается, а значит, подавляется активность АТФазы. Такой эффект полностью обратим и в опытах с лишенными сарколеммы волокнами. На ступенчатое повышение концентрации Са2+ от 10-7 до 10-5 М они реагируют постепенным увеличением силы сокращения и активности АТФазы, причем оба этих параметра достигают максимума при концентрации Са2+ 10-6-10-5 М.
Механизм активации ионами кальция мышечного волокна легче понять, рассмотрев структуру актиновых нитей (рис. 4). Каждый такой филамент длиной около 1 мкм и толщиной 5-7 нм состоит из двух закрученных одна вокруг другой цепочек мономеров актина толщиной 5 нм. Похожая структура получится, если взять две нити бус и скрутить их в виде спирали по 14 бусин в каждом витке
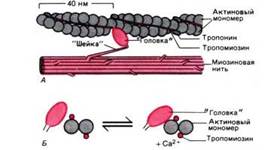
Рис. 4. Действие Cal+ во время активации миофибриллы. А. Актиновая и миозиновая нити на продольном сечении волокна. Б. Они же на его поперечном сечении. Когда Са2+ связывается с тропонином, тропомиозин попадает в желобок между двумя мономерами актина, обнажая участки прикрепления поперечных мостиков
Через регулярные промежутки примерно по 40 нм актиновые цепочки несут сферические молекулы тропонина, а в желобках между двумя цепочками лежат нити тропомиозина. Исследования с помощью рентгеноструктурного анализа (малоугловое рентгеновское рассеяние) показали, что в отсутствие Са2+, т. е. при расслабленном состоянии миофибрилл, длинные молекулы тропомиозина располагаются так, что блокируют прикрепление поперечных миозиновых мостиков к актиновым нитям. И напротив, под влиянием Са2+ молекулы тропомиозина глубже опускаются в желобки между цепочками мономеров актина, открывая участки прикрепления для поперечных мостиков. В результате те прикрепляются к актиновым нитям (рис. 4, Б), расщепляется АТФ и развивается мышечная сила.
Такой механизм активации обусловлен действием Са2+ на тропонин, который работает как "кальциевый переключатель": при связывании с Са2+ его молекула деформируется таким образом, что как бы заталкивает тропомиозин в желобок между двумя цепочками актиновых мономеров, т. е. в "активированное положение".
Хранение и высвобождение ионов кальция. Расслабленная мышца содержит более 1 мкмоль Са2+ на 1 г сырой массы. Если бы соли кальция не были изолированы в особых внутриклеточных хранилищах, обогащенные его ионами мышечные волокна находились бы в состоянии непрерывного сокращения.
Структура внутриклеточных систем хранения кальция в разных мышцах не вполне одинакова (скелетная мышца человека (рис. 1.; мышца лягушки-рис. 5). Во многих участках поверхностная мембрана мышечной клетки образует углубления в виде трубочек (диаметром 50 нм), перпендикулярных продольной оси волокна; эта система поперечных трубочек соединяется с внеклеточной средой и обычно окружает каждую миофибриллу на уровне Z-пластинок (у лягушки) или в области I-дисков (у высших позвоночных).
Перпендикулярно поперечным трубочкам, т. е. параллельно миофибриллам, расположена система продольных трубочек (истинный саркоплазматический ретикулум). Пузырьки на их концах (терминальные цистерны) прилегают к мембранам системы поперечных трубочек, образуя так называемые триады. В этих пузырьках и хранится внутриклеточный кальций. В отличие от поперечной системы продольная не сообщается с внеклеточной средой. Мембраны саркоплазматического ретикулума содержат работающий на энергии АТФ кальциевый насос, который осуществляет активный транспорт из миоплазмы в продольные трубочки, снижая таким образом примерно до 10-7М миоплазматическую концентрацию этих ионов в покоящейся (расслабленной) мышце.
Электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы внутрь клетки. При этом возбуждение быстро проникает в глубь волокна, переходит на продольную систему и в конечном счете вызывает высвобождение Са2+ из терминальных цистерн во внутриклеточную жидкость, окружающую миофибриллы, что и ведет к сокращению
При одиночном импульсе сокращение кратковременно расслабление мышцы вызывается обратным переносом активирующих ионов Са2+ посредством кальциевого насоса в каналы саркоплазматического ретикулума. Удаление ионов Ca2+ из миоплазмы идет до тех пор, пока их концентрация в ней не упадет до примерно 10-7 М. При этом подавляются активность АТФазы миозина и взаимодействие между актином и поперечными мостиками, которые отделяются от актиновых нитей.
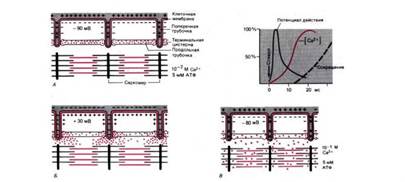
Рис. 5. Схема электромеханического сопряжения. А. Расслабленное мышечное волокно с поляризованной клеточной мембраной. Концентрация Ca2+ в нем ниже 10-7М. Б. Потенциал действия меняет полярность мембраны клетки и поперечных трубочек на противоположную; Ca2+ начинает выходить из терминальных цистерн. В. К моменту исчезновения потенциала действия внутриклеточная концентрация Ca2+ достигала примерно 10-5М, и саркомеры миофибрилл укоротились. Справа вверху: временная последовательность событий при электромеханическом сопряжении от "латентного" периода до начала сокращения (портняжная мышца лягушки при 0°С)
Распространение возбуждения вглубь волокна.
Этот процесс составляет первый этап электромеханического сопряжения (рис. 6). Воздействуя через микроэлектрод слабыми импульсами тока на мышечное волокно лягушки, эти авторы вызывали локальную деполяризацию такого маленького участка плазматической мембраны, что стимулировалась только одна поперечная трубочка (на уровне Z-пластинки). Возникающее в результате местное сокращение (контрактура) ограничивалось саркомерами поверхностных миофибрилл, непосредственно прилегающих к этой трубочке. По мере усиления стимула активировались все глубже расположенные миофибриллы. Очевидно, мембраны поперечных трубочек легко возбуждаются электрическим током, способны проводить возбуждение и составляют важное звено в процессе передачи сигнала от клеточной мембраны к хранилищам кальция.
Только за счет такой электрической передачи по поперечной системе возможна быстрая мобилизация запасов кальция в глубине волокна, и только этим можно объяснить очень короткий латентный период между стимулом и сокращением. Диффузия Ca2+ от поверхностной мембраны к миофибриллам, находящимся в центре мышечного волокна толщиной 100 мкм, продолжалась бы гораздо дольше, так что для волокон скелетных мышц подобный механизм можно исключить уже по временным соображениям.
Высвобождение кальция при одиночном сокращении. Блинке с коллегами выделили из светящихся медуз белок экворин, который при взаимодействии с Ca2+ излучает свет. После инъекции этого белка изолированное мышечное волокно закрепляли изометрически и раздражали электрическим током с интервалами 100 или 200 мс. С помощью высокочувствительного фотометра (фотоумножителя) регистрировалась люминесценция (излучение света) экворина, сопровождавшая внутриклеточное высвобождение Ca2+ (рис. 7). При стимуляции с частотой 5 Гц она была кратковременной, поскольку ионный насос вскоре перекачивал высвобожденный в миоплазму Ca2+ обратно в саркоплазматический ретикулум; при таком режиме мышца совершает одиночные сокращения. Однако при ритмичном раздражении с частотой 10 Гц (второй стимул поступает уже через 100 мс после первого) волокно расслабляется не полностью. Второе сокращение накладывается на остаточное сокращение после первого стимула, третье - на предыдущие и т. д.
Суммация одиночных сокращений ведет к росту как максимального напряжения в сократительном цикле, так и остаточной величины одиночных сокращений, хотя внутриклеточный уровень Ca2+ после каждого из них (судя по люминесценции) почти возвращается к уровню покоя.
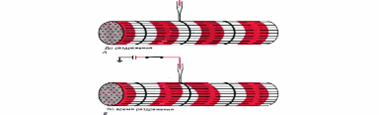
Рис. 6. Опыт, демонстрирующий возможность локальной активации Т-системы. Слабое локальное раздражение микрокатодом волокна поперечнополосатой мышцы лягушки (в области Z-пластинки, непосредственно над Т-трубочкой) вызывает укорочение прилегающих I-дисков: А - до, Б - во время раздражения
При этом опыт, представленный на рис. 7, показывает, что увеличение общего напряжения при стимуляции с интервалами по 100 мс нельзя объяснить повышением уровня внутриклеточного Ca2+.
Высвобождение Са2+ при тетанусе. Если стимулы поступают с высокой частотой (не менее 20 Гц), уровень Са2+ в интервалах между ними остается высоким, потому что кальциевый насос не успевает вернуть все ионы в продольную систему саркоплазматического ретикулума. Как показывает рис. 7, в таких условиях отдельные сокращения почти полностью сливаются. Это состояние устойчивого сокращения, или тетанус, наблюдается в том случае, когда промежутки между стимулами (или потенциалами действия в клеточной мембране) меньше примерно 1/3 длительности каждого из одиночных сокращений. Следовательно, частота стимуляции, необходимая для их слияния, тем ниже, чем больше их длительность; по этой причине она зависит от температуры. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, который приблизительно соответствует длительности потенциала действия.
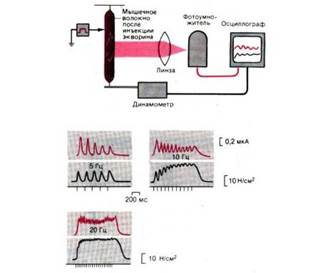
Рис. 7. Опыт, демонстрирующий внутриклеточное высвобождение Са2+ в мышечных волокнах. Люминесценция (красные кривые) и развитие изометрического напряжения в изолированном мышечном волокне шпорцевой лягушки при прямом раздражении импульсами тока длительностью по 0,5 мс с частотой 5, 10 и 20 Гц (моменты раздражения показаны штрихами под кривыми). Заметны суммация и слияние одиночных сокращений до (зубчатого) тетануса при повышении частоты стимуляции. Изометрическое напряжение измерено в ньютонах на 1 см2 площади поперечного сечения мышцы, а люминесценция, вызванная Ca2+, в единицах силы анодного тока фотоумножителя. Вверху: схема экспериментальной установки, использованной Блинксом и др.
Закон "все или ничего". Этому закону подчиняются быстрые волокна скелетных мышц.
Подпороговые стимулы не вызывают потенциалов действия и высвобождения Ca2+. Как только интенсивность стимула превысит определенный пороговый уровень, генерируется распространяющийся потенциал действия и происходит максимальное высвобождение Ca2+, это обеспечивает максимальную силу сокращения, уже не возрастающую при повышении интенсивности стимула.
Вместе с тем при электрическом раздражении целой мышцы сила ее сокращения зависит от интенсивности стимула. Например, если он едва превышает пороговый уровень, ответ по типу "все или ничего" наблюдается только в волокнах, находящихся вблизи от электрода, где плотность тока максимальна; для возбуждения всех волокон требуется гораздо более сильный (максимальный) стимул. Таким образом, только сверхмаксимальное раздражение может равномерно и достаточно надежно активировать изолированную целую мышцу.
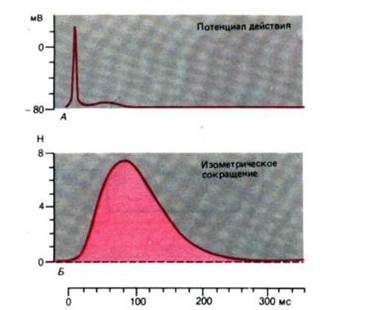
Рис. 8. Временной ход потенциала действия и изометрического сокращения поперечнополосатой мышцы (приводящей большой палец)
Мышечный изометрический поперечнополосатый миофибрилла
Закон "все или ничего" не означает, что ответ раздражаемого мышечного волокна будет всегда одинаков по величине. Например, если мышца только что расслабилась после тетануса, одиночный стимул часто вызывает гораздо более сильное одиночное сокращение, чем до такого "кондиционирования". Причины этой посттетанической потенциации известны так же мало, как и механизм мышечною утомления-снижения силы сокращения при ритмической стимуляции. В обоих случаях у потенциалов действия нормальная амплитуда. При кислородной недостаточности и в еще большей степени при нарушении метаболизма иодацетатом ритмическая стимуляция сопровождается не только снижением силы сокращения, но и замедлением расслабления; в конечном итоге, когда запас АТФ истощается, такая "отравленная" мышца вообще утрачивает способность к расслаблению--становится ригидной. Состояние необратимой ригидности и тетанус следует отличать от различных видов длительного напряжения, обсуждаемых ниже.
Похожие статьи
-
Блокирование нмс 1 уровень - пресинаптический - а) можно заблокировать ПД по мембране, блокаторами натриевых, калиевых каналов: тетродотоксин,...
-
Введение - Биофизика мышечного сокращения
Физика и биология, на первый взгляд, довольно далекие друг от друга науки. Но это только на первый взгляд. В действительности же в этих науках есть много...
-
Присоединение Са 2+ к тропонину устраняет постоянно существующее препятствие для взаимодействия поперечных мостиков с актином. Из результатов...
-
Молекулярный механизм сокращения - Биофизика мышечного сокращения
Один грамм ткани скелетной мышцы содержит примерно 100 мг "сократительных белков" - актина и миозина. Механизм их взаимодействия во время элементарного...
-
В естественных условиях одиночные сокращения скелетных мышц не встречаются. Происходит сложение, или суперпозиция, сокращений отдельных нейромоторных...
-
1) [-]продолговатый мозг 2) [-]промежуточный мозг 3) [+]большие полушария переднего мозга, мозжечок 4) [-]средний мозг Из какого белка состоит оперенье...
-
Одиночное мышечное сокращение возникает в ответ на одиночную стимуляцию Различают периоды 1 латентный период - с момента начала стимуляции до момента...
-
Строение скелетных мышц Каждая мышца состоит из параллельных пучков поперечнополосатых мышечных волокон. Каждый пучок одет оболочкой. И вся мышца снаружи...
-
Спинальные регуляции моторных функций - Принципы регуляции моторных функций
Систему регуляции моторных функций можно рассматривать как двухуровневую, где нижний спинальный уровень базируется на стреч-рефлексе - рефлексе на...
-
Скелет мышца кость позвоночник Инфекционные воспалительные заболевания мышц характеризуются мышечной слабостью в сочетании с диареей и снижением массы...
-
В гладкой мышце присутствуют 2 типа сопряжения между процессами возбуждения и сокращения - электромеханический тип и фармомеханисеский тип, возникающий...
-
Патологическая гистология сердечной мышечной ткани - Характеристика сердечной мышечной ткани
Различные повреждающие воздействия на сердце (прекращение притока артериальной крови, травмы, воспаления и др.) могут вызвать некроз мышечной ткани, т....
-
Мышечная ткань - Анатомия, конспект лекций
Охарактеризовать мышечную ткань, как ткань, обладающую способностью к сокращению, что позволяет осуществлять двигательные процессы в организме. Отметить...
-
Формирование скелетных мышц происходит на очень ранних этапах развития. На восьмой неделе внутриутробного развития различимы уже все мышцы, а к десятой...
-
Регуляция двигательных автоматизмов происходит с участием и стриопаллидарной системы. Показано, что стриарное тело функционально неоднородно: раздражение...
-
Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием белка миоглобина, который способен связывать...
-
+ возбудимые ткани - соединительные ткани - хрящевые ткани - костные ткани - эпителиальные ткани Специфичный лишь для данного вида ткани ответ на...
-
Регуляция секреции и выделение желчи - Состав желчи. Физиологическая желтуха новорожденных
Секреция желчи происходит непрерывно, независимо от того, находится ли пища в пищеварительном тракте или нет. Акт еды рефлекторно усиливает отделение...
-
Воспалительные заболевания мышц (миозиты) - общее название патогенетически разнородных заболеваний мышц, характеризующихся нарушением сократительной...
-
Лимфатическая система, Мышечная система, Костная система - Бронхиальная астма
Лимфатические узлы Результаты пальпации Околоушные Не пальпируются Подчелюстные Пальпируются, незначительно увеличены, эластической консистенции,...
-
Сокращаясь, мышцы выполняют работу. Работа зависит от их силы. Мышца тем сильнее, чем больше в ней мышечных волокон, то есть чем она толще. При пересчете...
-
Скелетный мышечный двигательный Развитие различных групп мышц происходит неравномерно. В 4-5 лет более развиты мышцы предплечья, отстают в развитии мышцы...
-
Костная система является очень прочной. Кость способна выдерживать большие нагрузки при ее сжатии и изломе. Основными составляющими кости являются...
-
Мышечная дистрофия - одно из самых тяжелых заболеваний опронодвигательного аппарата. В настоящее время нет лекарства на основе стволовых клеток для...
-
Регуляция дыхательных движений - Физиология и гигиена органов дыхания у детей
Регуляция дыхания осуществляется центральной нервной системой, специальные области которой обусловливают Автоматическое дыхание - чередование вдоха и...
-
Соединение костей, Мышечная система - Анатомия, конспект лекций
Объяснить, что в скелете человека в результате сочленения костей образуются либо механически более прочные непрерывные соединения, либо подвижные...
-
Мозжечок и регуляция двигательных функций - Принципы регуляции моторных функций
Важную роль среди надсегментарных структур, обеспечивающих управление движениями, играет мозжечок. Известно, что поражения мозжечка вызывают...
-
В процессе хемомеханического преобразования АТФ обеспечивает разъединение головки миозина и актинового филамента и энергетику для дальнейшего...
-
Строение внутренней оболочки сердца эндокарда - Характеристика сердечной мышечной ткани
Эндокард выстилает изнутри камеры сердца, папиллярные мышцы, сухожильные нити и клапаны сердца. Толщина эндокарда в различных участках неодинакова: толще...
-
Введение, Общая характеристика сердечной мышечной ткани - Характеристика сердечной мышечной ткани
В современной медицине все больший интерес вызывают вопросы лечения и профилактики заболеваний сердечно-сосудистой системы, возникновение которых в...
-
- миографии - реографии - определения диаметра мышц и нервов + определения порога раздражения - динамометрии К физиологическим свойствам возбудимых...
-
ФОБИИ. СТРЕСС - Регуляция физиологических процессов
Исследуются нейрохимические компоненты, характеризующие возникновение психических расстройств, определяемых в современной терминологии как "anxiety"...
-
К двигательным качествам относятся сила, скорость (быстрота), ловкость, гибкость и выносливость. Они обусловлены ростом и развитием всего организма....
-
Собственно саморегуляция представляет собой такой вариант управления, при котором отклонение какой-либо физиологической функции или характеристик...
-
Особенности нейроэндокринной регуляции стареющего организма - Возрастная биохимия
Возрастные изменения в гомеостазе пожилых людей многие авторы связывают с нарушениями, которые протекают в нейрогуморальных механизмах регуляции....
-
Железы смешанной секреции, Вывод - Гуморальная регуляция организма в онтогенезе
Яички и яичники - это половые органы, которые также относят к эндокринной системе, так как они вырабатывают гормоны, регулирующие половые функции. В...
-
Реализация функциональной активности фагоцитирующих клеток находится, в том числе, под контролем различных отделов нервной системы и регуляторных...
-
Строение скелетных мышц, Работа мышц - Анатомия, конспект лекций
Рассказать о строении скелетной мышцы, структурно-функциональной единицей которой является поперечнополосатое мышечное волокно. В мышце волокна...
-
Регуляция секреции гормонов - Понятие гормонов
Гормональная регуляция, регуляция жизнедеятельности организма животных и человека, осуществляемая при участии поступающих в кровь гормонов; одна из...
-
Заключение - Характеристика сердечной мышечной ткани
Анализ структурно-функциональных особенностей сердечной мышечной ткани показал, что, несмотря на то, что миокардиальная ткань состоит из отдельных...
Регуляция мышечного сокращения - Биофизика мышечного сокращения