Генетические детерминанты токсинов - Характеристика пестицидных белков Bacillus thuringiensis и их генетических детерминант
Уже показанное в разделе 3.1 многообразие как групп токсинов, так и индивидуальных токсинов является следствием высокой генетической пластичности Bacillus thuringiensis (Schnepf et al., 1998). Большинство генов, детерминирующих синтез токсинов, локализованы в больших плазмидах (Berry et al., 2002; Agaisse, Lereclus, 1995), многие из которых являются конъюгативными в природе (Schnepf et al., 1998). Помимо этого, в составе генома различных штаммов BT присутствуют разнообразные транслоцируемые генетические элементы - IS-элементы и транспозоны, поэтому Cry-гены в составе плазмид нередко входят в состав сложных транслоцируемых генетических конструкций (Добрица и др., 2001; Baum, 1994). В качестве примера можно привести ген Cry1А, фланкированный с обеих сторон инвертированными повторами, которые были определены как вставочные последовательности IS231 и IS232. Эти IS-элементы, очевидно, обеспечивают генетическую мобильность Cry1А гена, образуя вместе с ним типичный сложный транспозон (Schnepf et al., 1998). Кроме конъюгации, перенос генетического материала может обеспечиваться и путем трансдукции.
Все описанные выше особенности организации генома BT обеспечивают возможность обмена генетическим материалом между штаммами ВТ и другими видами бактерий, как близкородственными (B. cereus, B. anthracis), так и более отдаленными в генетическом отношении, такими, как B. subtilis, B. megaterium, B. sphaericus (Добрица и др., 2001; Han et al., 2006). Таким образом, для BT характерно участие в горизонтальном переносе генетического материала между видами семейства Bacillaceae, особенно с близкородственными видами, например, B. cereus (Schnepf et al., 1998).
Размер генома различных штаммов ВТ варьирует от 2,4 до 5,7 миллионов п. н. Для большинства штаммов ВТ размер плазмид, находящихся в бактериальной клетке, составляет 2 - 600 т. п.н. Cry-гены в большинстве своем локализованы в крупных плазмидах, в качестве примера можно привести мегаплазмиду pBtoxis, являющуюся генетическим детерминантом токсинов в клетках B. thuringiensis Subsp. israelensis (Berry et al., 2002). Размер этой плазмиды составляет 127 923 п. н., и она реплицируется по тета-механизму типа А. Плазмида pBtoxis кодирует все шесть токсинов, обнаруженных в клетках этой бактерии: Cry4Aa, Cry4Ba, Cry10Aa, Cry11Aa, Cyt1Aa и Cyt2Ba, а также несет несколько вставочных последовательностей и кодирует два протеина (P19 и P20), которые способствуют формированию кристаллических внутриклеточных включений и повышают жизнеспособность бактериальной клетки, действуя, вероятно, как шапероны (Berry et al., 2002). Помимо этого, в составе оперона, расположенного непосредственно вблизи точки инициации репликации, идентифицированы два гена, pBt156 и pBt157, кодирующие белки, необходимые для репликации плазмиды, массой 54,4 кДа и 11,8 кДа соответственно (Tang et al., 2006).
Некоторые общие характеристики pBtoxis даны в таблице 1, а комбинированная круговая карта плазмиды - на рис.6.
Таблица 1 Некоторые характеристики pBtoxis (По Berry et al., 2002)
|
Характеристика |
Значение |
|
Общий размер |
127 923 п. н. |
|
Содержание G + C |
32,42% |
|
Число кодирующих последовательностей |
125 |
|
Число псевдогенов |
8 |
|
Кодирующая плотность |
63,5% |
|
Средняя длина гена |
725 п. н. |
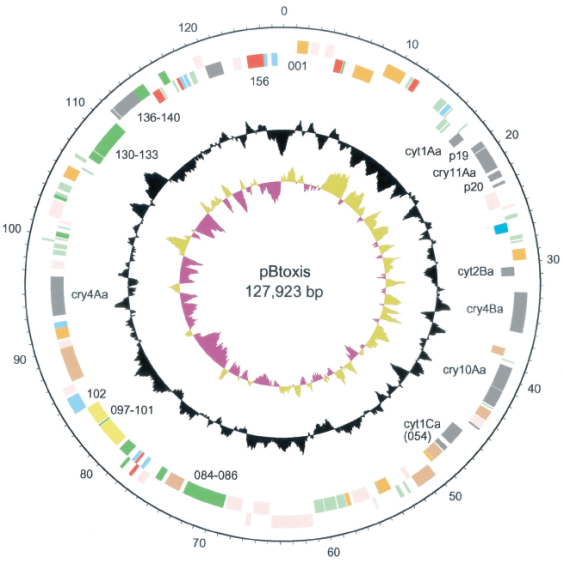
Рис.6. Комбинированная круговая карта мегаплазмиды pBtoxis
Внутренний круг отражает соотношение G-C / G+C (положительные значения - цвета хаки, отрицательные - пурпурного цвета), второй круг отражает содержание G + С. Два внешних круга дают представление о взаиморасположении генов на комплементарных нитях ДНК, при этом использованы следующие цветовые обозначения: серый - токсины и антибиотики, розовый - гены, связанные с транспозонами, оранжевый - гипотетически консервативные последовательности, красный - гены метаболизма ДНК, голубой - регуляторные гены, светло-зеленый - гены, детерминирующие поверхностные структуры клетки, бледно-зеленый - неизвестные гены, желтый - различные гены метаболизма. Внешняя шкала маркирована в т. п.н. (По Berry et al., 2002)
Cry-гены в зависимости от времени и механизма активации их экспрессии подразделяют на гены, зависящие от споруляции, и гены, не зависящие от споруляции. Протекание различных этапов процесса споруляции обеспечивается синтезом определенных мРНК и их трансляцией, причем последовательность синтеза задается тем, какой именно у-фактор свяжется с кор-ферментом РНК-полимеразы. Соответственно на разных стадиях споруляции последовательно активируются определенные у-факторы. Так, у Bacillus subtilis Процесс прохождения различных этапов жизненного цикла, включая споруляцию, регулируют шесть последовательно активируемых у - факторов: , , , , и. Такой же механизм активации экспрессии генов во время споруляции характерен и для BT, а ее соответствующие у-факторы обладают высокой степенью сходства по аминокислотной последовательности с аналогичными у-факторами B. subtilis (Agaisse, Lereclus, 1995).
Типичным примером Cry-гена, зависимого от споруляции, может служить ген Cry1A, транскрипция которого идет с двух сильных перекрывающихся промоторов, BtI и BtII, используемых последовательно, причем в транскрипции этого гена in vivo принимают участие оба специфических у-фактора, и (Adams et al., 1991; Agaisse, Lereclus, 1995).
К генам, не зависимым от споруляции, относится ген Cry3A, транскрипция с промотора которого идет слабо, но стабильно во время вегетативной фазы роста, активируется в конце экспоненциального роста и идет только до определенного этапа споруляции. В отличие от промоторов BtI и BtII, Cry3A промотор подобен промоторам, распознаваемым первичными у-факторами вегетативных бактериальных клеток, как, например, . Более того, экспрессия гена Cry3A усилена и пролонгирована в мутантных штаммах BT, не способных к инициации споруляции (Agaisse, Lereclus, 1995).
Данные о нуклеотидной последовательности промоторов некоторых Cry-генов приведены в таблице 2.
Таблица 2 Нуклеотидная последовательность промоторов некоторых Cry-генов (По Agaisse, Lereclus, 1995)
|
Промотор |
Область -35 |
Спейсер |
Область -10 |
|
Распознаваемый (B. thuringiensis) |
GCATNT |
N14 или 15 |
CATANNNT |
|
Cry1A(a) BtI |
GCATTT |
N15 |
CATATGTTT |
|
Распознаваемый (B. thuringiensis) |
Н/О |
Н/О |
TNATANNNTG |
|
Cry1A(a) BtII |
Н/О |
Н/О |
TCATAAGATG |
|
Распознаваемый (B. subtilis) |
TTGACA |
N17 или 18 |
TATAAT |
|
Cry3A |
TTGCAA |
N18 |
TAAGCT |
|
Примечание: Н/О - не определено. |
Похожие статьи
-
Общая характеристика и структура токсинов Токсины, продуцируемые BT, представляют собой белки с молекулярной массой от 130 до 140 кДа либо около 70 кДа...
-
Bacillus thuringiensis (BТ) относится к грамположительным аэробным спорообразующим эубактериям палочковидной формы и принадлежит к сем. Bacillaceae . Это...
-
Клетками-мишенями для белков Cry-семейства (т. н. д-эндотоксинов) являются эпителиальные клетки средней кишки насекомых, а также некоторых других...
-
Введение - Характеристика пестицидных белков Bacillus thuringiensis и их генетических детерминант
Объектом данной курсовой работы является Bacillus thuringiensis , а предметом - использование данного вида эубактерий как продуцента хозяйственно...
-
К чистым факторам переноса относят, например, факторы F и F-подобные (pAP22-4, pAP38, pAP39, pAP41) и pTRA1, фактор T, идентифицированный в E. coli,...
-
Этот оперон содержит гены, детерминирующие устойчивость к антибиотикам, а также в его состав может входить транспозон или его часть -...
-
Морская свинка обладает 32-мя парами хромосом. Таким образом получаем что общее число хромосом у морской свинки 64 шт о своему генотипу морские свинки...
-
Благодаря постоянному и беспорядочному броуновскому движению молекул воды [4] две идентичные по всем параметрам биомолекулы (БМ) способны не только...
-
Присутствие мобильных элементов в геноме является необходимым для генерирования генетического разнообразия посредством гомологической рекомбинации в...
-
Механизм, благодаря которому генетическая информация ДНК "транскрибируется" в матричную РНК, а затем транслируется в белок, выяснился через несколько лет...
-
- создают онкотическое давление крови, от которого зависит обмен воды между кровью и межклеточной жидкостью; - определяют вязкость крови, что в свою...
-
Первичная структура - понятие, обозначающее последовательность аминокислотных остатков в белке Пептидная связь - основной вид связи, определяющий...
-
Непременным компонентом вирусной частицы является какая-либо одна из двух нуклеиновых кислот, белок и зольные элементы. Эти три компонента являются...
-
Направления эволюции, Генетические и онтогенетические основы эволюции - Механизмы эволюции
Направление эволюции каждой систематической группы определяется взаимоотношениями между особенностями среды, в которой протекает эволюция данного...
-
Характеристики информации - Секреты молекулярной информации
Молекулярная биологическая информация мало чем отличается от других видов кодируемой информации. Ясно, что она выделяется чрезвычайно высокой плотностью...
-
Вирусы как независимые генетические системы Какое место занимают вирусы в биологическом мире? Каково их происхождение и кто их ближайшие родственники?...
-
Вирусная генетическая информация в трансформированных клетках - Строгие постулаты Коха
Все трансформированные вирусом клетки содержат его генетический материал. За исключением ДНК вируса ЭБ, который поддерживается в трансформированных им...
-
Генетический код - Великие открытия в генетике ХХ века
РНК передает инструкции от ДНК для создания белка. Но каков генетический код - последовательность инструкций, которая делать этот процесс возможным? В...
-
Гормоны (др.-греч. ?смЬщ -- возбуждаю, побуждаю) -- биологически активные вещества органической природы, вырабатывающиеся в специализированных...
-
Генетические методы изучения биопленок - Использование микробных биопленок в промышленности
Для выявления генов, участвующих в генетическом контроле любого процесса, используются методы направленного и ненаправленного (инсерционного)...
-
Заключение - Биосинтез белков. Ген и его роль в синтезе белков
В ходе написания курсовой работы мне удалось выяснить что биосинтез белков осуществляется во всех клетках про - и эукариот. Информация о первичной...
-
Этапы биосинтеза белка, Транскрипция - Биосинтез белков. Ген и его роль в синтезе белков
Транскрипция Мост между геном (кодонами) и белком обеспечивается РНК. Точнее, информация, закодированная в последовательности азотистых оснований ДНК,...
-
В цитозоле клеток 20 различных аминокислот присоединяются б-карбоксильной группой к 3'-гидроксильному акцепторному концу соответствующих тРНК с...
-
Ген. Взаимосвязь между геном и признаком - Биосинтез белков. Ген и его роль в синтезе белков
Долгое время ген рассматривали как минимальную часть наследственного материала (генома), обеспечивающую развитие определенного признака у организмов...
-
Введение - Биосинтез белков. Ген и его роль в синтезе белков
Способность клеток поддерживать высокую упорядоченность своей организации в хаотичной Вселенной зависит от генетической информации, которая реализуется,...
-
Расшифровка генетической информации - ДНК
Полимерные цепи белков состоят из мономерных звеньев - аминокислот и последовательность расположения их в белковой молекуле строго специфична. В связи с...
-
Различают два основных класса подвижных элементов: Транспозоны и Ретротранспозоны. Такая классификация основана на молекулярных механизмах, с помощью...
-
ВИЧ. СПИД. Профилактика СПИДа - Особенности генетического аппарата вирусов
ВИЧ - это вирус иммунодефицита человека, который вызывает тяжелое неизлечимое заболевание - синдром приобретенного иммунодефицита (СПИД). СПИД - это...
-
Характеристика пептидной связи - Первичная структура белков
Пептидная связь имеет характеристику частично двойной связи, поэтому она короче, чем остальные связи пептидного остова, и вследствие этого мало подвижна....
-
Введение - Особенности генетического аппарата вирусов
Цель: изучение генетического материала внеклеточных организмов. Задания: 1. Многоуровневая организация генома. 2. Геном РНК - вирусов. В настоящее время...
-
Классификация простейших, Характеристика класса - Морфология и систематика простейших
Медицинское значение имеют простейшие относящиеся к классам: Саркодовые; Жгутиковые; Споровики; Инфузории. Характеристика класса Саркодовые:...
-
Вирусы являются генетическими внутриклеточными паразитами, способными к размножению только в живых системах. Вирусы как облигатные внутриклеточные...
-
Механизм дробления - Дробление, его характеристика у разных животных. Типы бластул
Клеточный цикл соматических клеток делится на четыре стадии (фазы). За митозом (М) следует пресинтетический период (G1), после которого происходит синтез...
-
Типы бластул - Дробление, его характеристика у разных животных. Типы бластул
Имеется 5 основных типов бластул: - Целобластула. - Бластоциста. - Амфибластула. - Дискобластула. - Перибластула. Целобластула характерна для иглокожих и...
-
Общая характеристика эндокр. желез - Общая анатомия человека
Эндокринология - наука о железах внутренней секреции. Эндокринные железы не имеют выводных протоков и выделяют вырабатываемый или секрет -- гормоны (от...
-
Анализаторы. Общая характеристика и строение - Основные этапы индивидуального развития человека
Сенсорная (чувствительная) информация играет очень важную роль в жизнедеятельности человека. Она поступает в нервную систему различными путями. Через...
-
Зарождение иммунологии - Характеристика основ иммунологии
Начало развития иммунологии относится к концу XVIII века и связано с именем Э. Дженнера, впервые применившего на основании лишь практических наблюдений...
-
Введение - Характеристика основ иммунологии
Мечников открыл первую клетку иммунной системы, которую назвали фагоцит, или макрофаг. Греческое слово "фаг" означает поедание, пожирание. Фагоцитоз был...
-
Бластула - Дробление, его характеристика у разных животных. Типы бластул
У многих яйцеклеток еще на ранних стадиях дробления внутренние концы бластомеров расходятся, и между ними возникает сначала небольшая, а затем все...
-
Введение - Дробление, его характеристика у разных животных. Типы бластул
Оплодотворение играет чрезвычайно важную роль в развитии, но оно лишь только первая его ступень. Зигота с ее новым генетическим потенциалом и новым...
Генетические детерминанты токсинов - Характеристика пестицидных белков Bacillus thuringiensis и их генетических детерминант