Классификация подвижных генетических элементов, их структура и способы перемещения - Мобильные генетические элементы эукариот: транспозоны и ретротранспозоны
Различают два основных класса подвижных элементов: Транспозоны и Ретротранспозоны. Такая классификация основана на молекулярных механизмах, с помощью которых перемещаются подвижные элементы.
Транспозоны перемещаются с участием комплекса белков, обеспечивающего активность фермента транспозазы, которая "узнает" элемент и обеспечивает его перенос на новое место. Мобильные элементы обычно фланкированы короткими повторами генома хозяина, служащими целевой последовательностью для их интеграции, причем они часто оканчиваются инвертированными повторами. Инвертированные повторы - это последовательности, направленные навстречу друг другу. Они необходимы для перемещения элемента, которое осуществляется благодаря их сближению друг с другом и узнаванию транспозазами. Инвертированные повторы сближаются и точно отрезаются от соседних участков ДНК хозяина [2,3].
Успешному вырезанию элемента способствует дополнительная сверхспирализация двухнитевой спирали ДНК, обеспечивающая изгибы двойной спирали и сближение отдельных ее участков. Вырезанный транспозон внедряется в район вносимого транспозазой разрыва в молекуле-мишени и сшивается с ДНК хозяина в новом месте (см. рис. 1). Разрыв и зашивание осуществляются транспозазой и вспомогательными белками. Транспозаза может кодироваться как самим подвижным элементом, который будет перемещаться, так и другой копией элемента, локализованной в том же геноме в отдалении.
![механизм перемещения днк-транспозона [4]](/images/image002-493.jpg)
Рис.1. Механизм перемещения ДНК-транспозона [4]
Брешь в ДНК, оставляемая после вырезания транспозона, может залечиваться - застраиваться с участием гомологичного участка, например сестринской, только что редуплицированной молекулы ДНК[2].
Число ДНК транспозонов не увеличивается при транспозиции, но число их копий в видовом геноме обычно может достигать от нескольких штук до нескольких сот. Эти элементы кодируют один или несколько белков, достаточных для того, чтобы транспозиция осуществилась. Структура их концевых последовательностей является более сложной, чем у ретротранспозонов, поскольку содержит два типа последовательностей, действующих в Cis - положении, таких, как концевые инвертированные повторы (КИП) и субконцевые последовательности, которые совместно регулируют механизм вырезания-сшивания [1].
Другой большой класс подвижных элементов - это Ретротранспозоны, они не вырезаются из хромосомы, как это делают транспозоны. Механизм их перемещения основан на существовании открытой в 1970 году Г. Теминым и Д. Балтимором реакции обратной транскрипции - синтеза нити ДНК на РНК. Химическая реакция протекает так же, как при образовании нити комплементарной ДНК на ДНК-матрице при репликации двухнитевой молекулы ДНК. Ретротранспозоны используют РНК в качестве посредника (рис.1) [3,5].
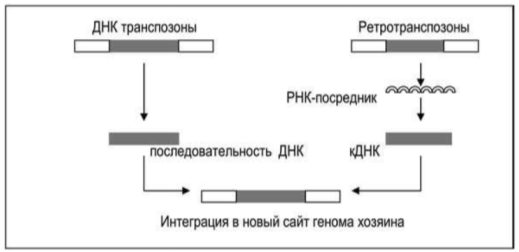
Рис.2. Tранспозиция в новый сайт генома хозяина двух типов мобильных элементов посредством вырезания и последующей интеграции участа ДНК (слева) и с помощью РНК-посредника, сопровождаемая обратной транскрипцией кДНК и ее вставкой в новый сайт генома (справа) [3]
Ретротранспозоны транскрибируются подобно другим последовательностям генома, и полученная РНК используется в качестве матрицы для синтеза кДНК с помощью фермента обратной траскриптазы, кодируемого ДНК транспозона [3]. Новосинтезированный ретротранспозон встраивается в другой участок генома.
Активные ретротранспозоны млекопитающих делятся на три основные семьи: Alu-повторы, ДДП-1, SVA.
ДДП-1 ретротранспозоны - длинные диспергированные повторы - тип ретротранспозонов, который широко распространен у млекопитающих и составляет до 20 % генома. ДДП-1 элементы имеют длину около 6 тысяч пар оснований. Большинство этих ретротранспозонов в геноме представлено неполно, хотя существует примерно 150 полных и потенциально мобильных ДДП-1 элементов в последовательности ДНК человека и примерно 3000 - у мыши [6].
ДКП - длинные концевые повторы - ретротранспозоны, имеющие конечные повторяющиеся последовательности, которые играют важную роль в транскрипции и обратной транскрипции РНК транспозона. ДКП-элементы кодируют белки Pol и Gag, которые близки к белкам ретровирусов, но, в отличие от последних, ДКП не хватает белков, которые смогли бы сформировать внешнюю оболочку (суперкапсид) и выйти из клетки.
КДП - короткие диспергированные повторы, являются неавтономными ретротранспозонами: они требуют активности ДДП-1 элементов для передвижения, в ДНК последовательности КДП содержат только участок связывания РНК-полимеразы. В число КДП входят Alu-ретротранспозоны [7].
Alu-Повтор (Alu от Arthrobacter luteus) -- широко распространенные мобильные элементы в геноме человека. Alu-элементы имеют длину около 300 пар оснований и часто расположены в интронах, участках генома, которые не транслируются, и межгенных участках. Приставку Alu- ретротранспозоны получили за то, что они содержат последовательность распознавания рестрикционного энзима AluI. Анализ последовательностей показал, что Alu - элементы возникли у приматов примерно 65 миллионов лет назад от гена 7SL РНК, который входит в рибосомный комплекс. Alu-ретротранспозоны не имеют собственной обратной транскриптазы, поэтому для передвижения им необходимые ферменты ДДП-1 элементов [8].
SVA - мобильные элементы длиной в 2-3 тысячи пар оснований ДНК, состоящие из нескольких частей. SVA элементы значительно варьируют в длину из-за разного количества составляющих повторов. Они не являются автономными и нуждаются в белках, закодированных в ДДП1 ретротранспозонах для передвижения, но они активны в геноме человека. SVA-элементы претерпевают высокий уровень метилирования ДНК в большинстве тканей человека[6].
Ретротранспозоны широко распространены у эукариот, населяя геномы дрожжей, растений, насекомых и позвоночных, включая человека. Ретротранспозоны пока не обнаружены у прокариот. Тысячи различных семейств ретротранспозонов могут занимать до 70-85% ядерной ДНК растений и животных. Около 75% генома кукурузы представлено ретротранспозонами, главным образом ДКП типа. В то же время маленький геном дрожжей Saccharomices cerevisiae содержит только лишь пять семейств ДКП ретротранспозонов, занимающих лишь 3% их геномной ДНК. В геноме Arabidopsis обнаружено несколько сот семейств ДКП и без ДКП ретротранспозонов, которые составляют около 14% его ядерной ДНК. У млекопитающих число LINE и SINE может достигать сотни тысяч копий, составляя 35% величины их генома. В целом разнообразие и численность мобильных элементов в геномах растений выше, чем у представителей других царств. На рис.2. отображена представленность различных типов транспозонов в геноме человека [1,2,3,9].
![представленность транспозонов в геноме человека [9]](/images/image004-697.png)
Рис.3. Представленность транспозонов в геноме человека [9]
Похожие статьи
-
Введение - Мобильные генетические элементы эукариот: транспозоны и ретротранспозоны
В начале 1950_х гг. Б. Мак-Клинток на основе генетических экспериментов, выполненных на кукурузе, постулировала наличие в геноме высших эукариот...
-
Сразу же отметим, что изменить положение своей БД для ответа на изменение внешней среды в водах с замедленным течением и тем более - в стоячих водах...
-
Геном пластид - Генетика пластид
В 1961 году X. Рис и В. Плаут обнаружили, что участок хлоропласта в одноклеточной водоросли хламидомонады окрашивается положительно по Фельгену. При...
-
Повторяющаяся ДНК - Геном человека
В любом сложном геноме ДНК можно подразделить на два типа: однокопийную ДНК (т. е. последовательности, представленные в гаплоидном геноме единственными...
-
Строение гена человека - Геном человека
В каждой диплоидной клетке человека 46 хромосом, содержащих около 6пг ДНК и 3,2 х 109 пар нуклеотидов, длина всех молекул ДНК около 2м. А если учесть,...
-
Генетический скрининг взрослых - Геном человека
В настоящее время нет общенациональных программ генетического скрининга взрослого населения, но некоторые достижения заслуживают упоминания. Лучшим...
-
Этот оперон содержит гены, детерминирующие устойчивость к антибиотикам, а также в его состав может входить транспозон или его часть -...
-
В работах [17, 18] уже говорилось о том, что по мере остывания молодой Земли, температура водных растворов и в зонах извержения подводных вулканов также...
-
Благодаря постоянному и беспорядочному броуновскому движению молекул воды [4] две идентичные по всем параметрам биомолекулы (БМ) способны не только...
-
Прогресс технологии генотипирования и разработка современных статистических методов предоставили новые возможности для изучения роли вариабельности...
-
Классификации основных форм поведения - Поведение животных
Поведение животных бесконечно разнообразно по своим формам, проявлениям и механизмам. В настоящее время накоплен большой материал, который характеризует...
-
Классификация мутаций - Мутация
1. По способу возникновения. Различают спонтанные и индуцированные мутации Спонтанные происходят в природе крайне редко с частотой 1-100 на миллион...
-
Вирусы как независимые генетические системы Какое место занимают вирусы в биологическом мире? Каково их происхождение и кто их ближайшие родственники?...
-
Вирусные заболевания. - Особенности генетического аппарата вирусов
Эволюция вирусов и вирусных инфекций. Хотя вирусы не являются полноценными живыми организмами, их эволюционное развитие имеет много общего с эволюцией...
-
ДНК-содержащие вирусы. - Особенности генетического аппарата вирусов
К ДНК-содержащим вирусам относятся многие вирусы бактерий - бактериофаги (или просто фаги). Некоторые мелкие фаги (например, фаг М13) при репродукции не...
-
В настоящее время оценка генетической структуры различных популяций осуществляется с использование биологических и небиологических источников информации,...
-
Циклы биогенных элементов - Структура Земли и ее циклы
Другой пример глобального цикла, также хорошо известный каждому со школьной скамьи, - цикл углерода, приведенный на рис. 11. Главный резервный фонд...
-
Определено расположение субъединиц в геномах вирусов, прокариотов и эукариотов, послужившее основой для создания универсального генетического кода....
-
Именно броуновское движение органических молекул в водной среде является причиной не только самоорганизации БС, но и их самовосстановления....
-
Когнитивные различия В среднем мужчины и женщины имеют одинаковые интеллектуальные способности. Это подтверждается результатами тестов на выявление...
-
Возможные последствия клонирования человека - Клонирование: достижения, перспективы, проблемы
Споры по поводу запрещения клонирования чуть было не привели к свертыванию проекта Вилмута, но ученым удалось отстоять результаты своей работы и...
-
Скорпионы, Общая характеристика, Яд скорпионов - Пауки и скорпионы
Общая характеристика Скорпионы издавна докучали людям своим ядовитым жалом. Они относятся к классу паукообразных и состоят в родстве с пауками и...
-
Отрицательные виды взаимоотношений между организмами - Взаимосвязи животных в природе
Существуя вместе на планете, животные могут не только помогать друг другу, но и причинять вред. Виды взаимоотношений, которые считаются отрицательными, в...
-
Синантропные и культурные растения - Синантропная флора Дальнего Засвияжья г. Ульяновска
С деятельностью человека связано появление синантропных растений (антропофитов). Появились синантропные растения непреднамеренно помимо его воли человека...
-
Обмен белков в организме человека - Строение, функции и значение белков
Важный критерий пищевой ценности белков - доступность аминокислот. Аминокислоты большинства животных белков полностью высвобождаются в процессе...
-
Понятие ген - Перспективная наука геномика
Структурной и функциональной единицей генома является ген. Ген контролирует развитие определенного признака или свойства организма. Совокупность генов...
-
Происхождение вирусов и происхождение клетки - Строгие постулаты Коха
Проблема происхождения вирусов - это, по существу проблема независимости генетических элементов в репродуктивном и эволюционном отношении. Основные...
-
Биопринтер -- биологическая вариация технологии reprap, устройство, способное из клеток создавать любой орган, нанося клетки слой за слоем, уже создано....
-
В наше время очень актуально проблема вирусов. Она привлекает внимание все большего числа ученых. Когда стало известно о существовании вирусов, никто и...
-
Возможные последствия клонирования человека - Клонирование
Споры по поводу запрещения клонирования чуть было не привели к свертыванию проекта Уилмута, но ученым удалось отстоять результаты своей работы и...
-
Замена аминокислоты на поверхности гемоглобина А - Первичная структура белков
В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 в-цепи, в которых глутамат, высокополярная отрицательно заряженная...
-
Утомление -- это состояние временного снижения работоспособности человека. Оно развивается вследствие напряженной или длительной умственной либо...
-
Вирусная генетическая информация в трансформированных клетках - Строгие постулаты Коха
Все трансформированные вирусом клетки содержат его генетический материал. За исключением ДНК вируса ЭБ, который поддерживается в трансформированных им...
-
В основе расходования воды растительным организмом лежит процесс испарения -- переход воды из жидкого в парообразное состояние, происходящий при...
-
Несовместимость и группы несовместимости - Молекулярная и генетическая организация плазмид
К одной группе несовместимости (inc-группа) относят плазмиды, которые несовместимы между собой, но совместимы с любой плазмидой из другой группы....
-
Гликозиды встречаются в коре, плодах, корнях, клубнях, цветках и других частях растений. Иногда в одном растении содержится несколько разных гликозидов....
-
Наследственная информация бактерий хранится в ДНК, которая в прокариотической клетке является циркулярно замкнутой, двух цепочечной, суперспирализованной...
-
Обычно гликозиды классифицируют по типу агликона. Основные классы гликозидов перечислены ниже. Тиольные гликозиды (тиоцианатные, изотиоцианатные, сульфо...
-
Онкогенные вирусы. классификация - Частная микробиология
РНК-содержащие: семейство Retroviridae. ДНК-содержащие: семейства Papillomaviridae, Polyomaviridae, Adenoviridae 12, 18, 31, Hepadnaviridae,...
-
Генетический код - Великие открытия в генетике ХХ века
РНК передает инструкции от ДНК для создания белка. Но каков генетический код - последовательность инструкций, которая делать этот процесс возможным? В...
Классификация подвижных генетических элементов, их структура и способы перемещения - Мобильные генетические элементы эукариот: транспозоны и ретротранспозоны