Особенности переваривания и всасывании жиров у детей - Обмен и функции липидов
Главная особенность переваривания жиров в раннем детском возрасте заключается в том, что примерно половина жиров расщепляется в желудке. Данная особенность обусловлена следующими обстоятельствами:
- 1. жиры молока находится в эмульгированном состоянии 2. при грудном вскармливании в переваривании жиров участвует липаза грудного молока 3. в процессе сосания у грудного ребенка вырабатывается лингвальная липаза, которая оказывает эффект в желудке 4. активно вырабатывается желудочная липаза с оптимумом рН около 5,0 5. у детей в желудке менее кислая среда, приближенная к оптимуму рН для липаз 6. активность панкреатической липазы у детей снижена 7. в детском возрасте менее активен синтез желчных кислот, повышена их потеря через кишечник и замедлена циркуляция.
Всасывание жиров у детей происходит с большей скоростью, чем у взрослых в связи с высокой проницаемостью слизистой кишечника.
Транспорт жиров кровью
Гидрофобные жиры не могут транспортироваться кровью самостоятельно. Они переносятся в следующих формах:
- 1. липопротеиды (липопротеины) - белково-липидные комплексы 2. хиломикроны - жировые капли, образующиеся в млечном соке 3. свободные жирные кислоты транспортируются в комплекте с альбуминами
Хиломикроны - это мельчайшие капельки жира с размерами около 500 нм, плотностью 0,95 г/см3, состоящие из 2% белка и 90% ТАГ. Хиломикроны синтезируется в слизистой кишечника, считаются транспортной формой пищевых (экзогенных) жиров организме. Хиломикроны попадают сначала в лимфу, а затем разносятся кровью в основном в жировые депо (>50%), а также к печени, легким, мышечной ткани.
Липопротеиды (ЛП) являются основной транспортной формой жиров.
По электрофоретической подвижности различают: пре в - ЛП, в - ЛП, б - ЛП
По плотности выделяют:
- - ЛП очень низкой плотности (ЛПОНП) - ЛП низкой плотности (ЛПНП) - ЛП высокой плотности (ЛПВП) - ЛП промежуточной плотности - ЛП очень высокой плотности
Все ЛП построены по общему принципу. В центре частицы находится гидрофобное ядро, в которое входят ТАГ и эфиры холестерина, вокруг него формируется гидрофильная оболочка, в которую входят ФЛ, холестерин. На поверхности располагаются белки - апопопротеины (АроPt).
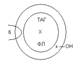
Различают несколько видов АроPt: A, B, C, E. Они формируют структуру липопротеидных частиц, взаимодействуют с тканевыми рецепторами к ЛП, являются активаторами ферментов обмена ЛП
ЛП осуществляют транспорт липидов, жирорастворимых витаминов и гидрофобных гормонов.
Закономерности строения липопротеидов в ряду: ЛПОНП >ЛПНП>ЛПВП представлены в таблице.
Таблица 1
|
ЛПОНП |
ЛПНП |
ЛПВП | |
|
Размеры уменьшаются |
80 нм |
25 нм |
10 нм |
|
Плотность возрастает |
1,006 г/см3 |
1,006-1,06 г/см3 |
1,25 г/см3 |
|
% содержания белков увеличивается |
10% |
25% |
50-60% |
|
% содержания липидов уменьшается |
60% ТАГ |
45-48% холестерин |
|
ЛПОНП - синтезируется в печени, считаются основной транспортной формой эндогенных жиров. В эндотелии сосудов ЛПОНП и хиломикроны подвергаются действию фермента липопротеидной липазы, которая расщепляет в их составе ТАГ. В результате в составе ЛП повышается доля холестерина, и ЛПОНП превращаются в ЛПНП.
ЛПНП считаются транспортной формой холестерина от печени к органам и тканям. В тканях имеются рецепторы и ЛПНП, при участии которых происходит поглощение холестерина с последующим использованием его на построение мембран, синтез стероидов, депонированием в виде эфиров.
ЛПВП синтезируется в печени в виде дисковидных структур. Они считается транспортной формой холестерина из тканей к печени. В кровотоке при контакте с эндотелием происходит поглощение холестерина ЛПВП. Они постепенно превращаются в сферические структуры и переносят холестерин в печень. В поглощении холестерина частицами ЛПВП участвует фермент ЛХАТ (лицитинхолестеролацилтрансфераза), который в составе ЛПВП переносит остатки жирных кислот с фосфолипидов на холестерин с образованием эфиров холестерина. Эфиры холестерина более гидрофобны по сравнению со свободным холестерином и, в силу этого, погружаются внутрь ЛП частицы.
У детей общее содержание ЛП ниже, чем у взрослых. В детском возрасте снижена концентрация хиломикронов и ЛПОНП, повышено содержание ЛПВП, в которых повышено содержание гидрофильных компонентов.
Содержание липидов в крови взрослых:
Таблица 2
|
Общие липиды 4-8 г/л |
|
ТАГ 1-2,5 ммоль/л |
|
ФЛ 2,5-3,0 ммоль/л |
|
Холестерин 3,5-5,2 ммоль/л |
|
Свободные жирные кислоты 0,5 - 1,0 ммоль/л |
У детей содержание общих липидов ниже, чем у взрослых. В то же время концентрация свободных жирных кислот у детей повышено примерно в два раза.
Большая часть переносимых кровью липидов откладывается в жировых депо, к которым относятся подкожно-жировая клетчатка, большой и малый сальники. У детей наиболее активно депонирование жиров происходит в возрасте 1 года, 7 лет и в пубертатном периоде. В раннем детском возрасте у детей важным видом жировой ткани является бурая жировая ткань. Она локализована в основном на спине, на груди, имеет бурый оттенок, который обусловлен большим содержанием митохондрий и Fе - содержащих цитохромов. В бурой жировой ткани происходит нефосфолирирующее окисление жиров, которое сопровождается выделением тепловой энергии (она является органом термогенеза). Жировое депо у детей легко истощается при нарушении питания, болезнях, стрессе. Липиды в жировых депо постоянно обновляются.
Обмен триацилглицеринов
Распад триацилглицеринов в тканях (липолиз)
Триацилглицерины поэтапно расщепляется тканевыми липазами.

Ключевым ферментом липолиза является гормональнозависимая ТАГ-липаза. Образующиеся на этом этапе распада жиров глицерин и жирные кислоты окисляются в тканях с образованием энергии.
Окисление жирных кислот.
Различают несколько вариантов окисления жирных кислот: б - окисление, в - окисление, щ - окисление. Основным вариантом окисления жирных кислот является в - окисление. Оно наиболее активно протекает в жировой ткани, печени, почках и сердечной мышце.
В - окисление заключается в постепенном отщеплении от жирной кислоты двух углеродных атомов в виде ацетил - КоА с освобождением энергии. Запас жирных кислот сосредоточен в цитозоле, где протекает активация жирных кислот с образованием ацил - КоА
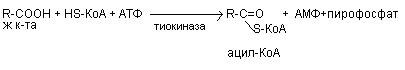
Последующее в-окисление ацил-КоА происходит в митохондриях. Митохондриальная мембрана непроницаема для длинноцепочечных ацил - КоА. В переносе их внутрь митохондрий участвует специальный переносчик карнитин (метил, гидропроизводное аминомасляной кислоты). Ацил - КоА образует с карнитином комплекс, который после переноса жирной кислоты внутрь митохондрий распадается.
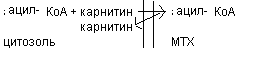
Химизм в - окисления насыщенных жирных кислот
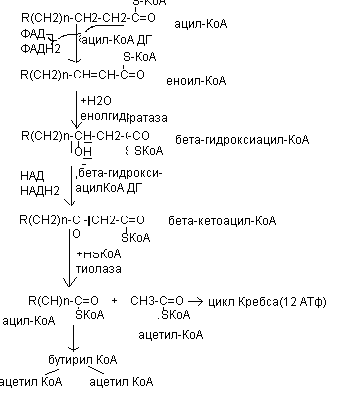
Энергетическая эффективность бета - окисления жирных кислот складывается из энергии окисления ацетил - КоА в цикле Кребса и энергии, освобождающейся в самом бета - цикле. Энергия окисления жирной кислоты тем выше, чем длиннее ее углеродная цепь. Количество молекул ацетил - КоА из данной жирной кислоты и количество образующихся из них молекул АТФ определяется по формулам:
N=N/2,
Где n - количество молекул ацетил - КоА,
N - число атомов углерода в жирной кислоте.
Количество молекул АТФ за счет окисления молекул ацетил-КоА = (N/2)*12
Число в - циклов окисления на один меньше, чем количество образующихся молекул ацетил-КоА, поскольку в последнем цикле масляная кислота за один цикл переходит в две молекулы ацетил-КоА, и рассчитывается по формуле
Количество в - циклов = (N/2)-1
Количество молекул АТФ в в - цикле рассчитывается, исходя из последующего окисления образовавшихся в нем НАДН2 (3 АТФ) и ФАДН2 (2 АТФ) по формуле
Количество молекул АТФ, образующихся в бета-циклах = ((N/2)-1)*5
2 макроэргические связи АТФ расходуются на активацию жирной кислоты
Суммарная формула для подсчета выхода АТФ при окислении насыщенной жирной кислоты имеет вид: 17(N/2)-7.
При окислении жирных кислот с нечетным числом углеродных атомов образуется сукцинил - КоА, который вступает в цикл Кребса.
Окисление ненасыщенных жирных кислот на начальных стадиях представляет обычное бета - окисление до места двойной связи. Если эта двойная связь находится в бета - положении, то продолжается окисление жирной кислоты со второго этапа (минуя стадию восстановления ФАД> ФАДН2). Если двойная связь находится не бета - положении, то ферментами еноилтрансферазами связь перемещается в бета - положение. Таким образом, при окислении ненасыщенных жирных кислот образуется меньше энергии по формуле (теряется образование ФАДН2):
7(N/2)-7-2m,
Где m-число двойных связей.
Похожие статьи
-
Всасывание продуктов расщепления жиров - Обмен и функции липидов
Всасывание продуктов расщепления жиров происходит в тонком кишечнике и определяется водорастворимостью или водонерастворимостью образовавшихся продуктов...
-
Спиртовой и жирнокислотный состав основных жиров тканей человека - Обмен и функции липидов
Липиды - неоднородные в химическом отношении органические вещества, нерастворимые в воде, но растворимые в органических растворителях. Жиры включают в...
-
Функции жиров - Обмен и функции липидов
- Энергетическая функция: снабжают организм энергией. Калорическая ценность жиров выше, чем у углеводов и белков (1г жира дает при окислении около 9...
-
Водораствроримые витамины Большинство водорастворимых витаминов, поступающих с пищей или синтезируемых кишечными бактериями, проявляют активность после...
-
Особенности азота - Свойства азота
У атома азота на один электрон больше, чем у атома углерода; согласно правилу Гунда, этот электрон занимает последнюю вакантную 2р-орбиталь. Атом азота в...
-
В 2005 году В. Б. Спиричевым предложена новая классификация витаминов, основанная на характере их специфических функций (функциональная классификация) В...
-
Витамины и их значение - Классификация и номенклатура витаминов. Роль витаминов в обмене веществ
Витамины - это низкомолекулярные природные органические биологически активные вещества, абсолютно необходимые для нормальной жизнедеятельности любого...
-
Используется адаптивная нейро-нечеткая система вывода ANFIS, функционально эквивалентная системе нечеткого вывода Сугено. Вывод осуществляется за два...
-
Аппроксимация функции предпочтения ЛПР нейронными сетями имеет в работе ту особенность, что процесс обучения нейронных сетей происходит в условиях малой...
-
Химические свойства - Свойства азота
Азот находиться в верхнем правом углу периодической системы, в котором сосредоточены неметаллы с наибольшими сродствами к электронам. Поэтому он должен...
-
Изменение жиров при хранении - Химия и физика молока
Современные представления о механизме окисления жиров. При неблагоприятных условиях хранения в жирах протекают различные процессы, отрицательно влияющие...
-
КАРОТИН КАК ПРОВИТАМИН А - Биологическая роль каротина и каротиноидов
Общие сведения Провитаминная сущность каротина заключается в том, что этот пигмент способен расщепляться в организме человека и многих животных с...
-
Флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД) функционируют как коферменты для широкого спектра окислительных ферментов и остаются связанными...
-
Функции, Предельная недостаточность - Витамин Вс (фолиевая кислота)
Тетрагидрофолиевая кислота, являющаяся активной формой фолата в организме, действует в качестве кофермента во многих базовых метаболических реакциях. Она...
-
МЕТОДЫ ОСАЖДЕНИЯ, ИОННОГО ОБМЕНА, МЕМБРАННЫЕ МЕТОДЫ - Химические свойства и строение воды
Современные технологии позволяют изготавливать объемные или плоские фильтрующие материалы с однородными каналами практически любого размера. Мембранный...
-
Аннотация - Точность математического прогнозирования как функция количества учитываемых факторов
В статье рассмотрена точность прогнозирования экономических показателей в зависимости от количества используемых параметров на основе математического...
-
ПРАВАЯ И ЛЕВАЯ ТРОЙКИ ВЕКТОРОВ Линейно независимые векторы, и образуют Правую Тройку векторов, если они имеют такую же ориентацию, как соответственно...
-
Элементы теории магнетизма. Доменная структура и петля гистерезиса (ферро, ферри-, антиферромагнетики). Важнейшие типы магнитомягких и магнитожестких...
-
Как и каждый достаточно ярко выраженный класс экономико-математических моделей, совокупность моделей календарного планирования обладает рядом...
-
Разберемся, что есть в грудном молоке изначально и не зависит от того, чем питается женщина, а чего может и не хватать, если рацион мамы однообразный и...
-
Особенности эконометрического метода Эконометрическая модель -- основное понятие эконометрии, экономико-математическая модель, параметры которой...
-
При использовании статистических методов прогнозирования во многих случаях необходимо знать возможную ошибку прогноза, т. е. тот интервал, в котором...
-
Особенности структуры аморфных наночастиц - Коллоидная химия
По геометрическому признаку (мерности дисперсных частиц) наносистемы можно разделить на три группы. 1. Трехмерные (объемные) наночастицы , у которых все...
-
Источники поступления в живой организм Суточную потребность в сере можно обеспечить правильно организованным рациональным питанием. Основными источниками...
-
Морфологические особенности состояний - Системная революция и принцип дуального управления
Состояние есть совокупность (система) переменных признаков, вполне характеризующих изменчивую часть образа системы. Поскольку реальная система имеет...
-
Проникновение математики в экономическую науку связано с преодолением значительных трудностей. В этом отчасти была "по-винна" математика, развивающаяся...
-
Возраст Чувствительность к лекарственным средствам меняется в зависимости от возраста. В связи с этим выделилась так называемая перинатальная...
-
К числу таких реакций относятся взаимодействия солей двухвалентных катионов (кроме Ca2+, Sr2+, Ba2+) с водными растворами карбонатов натрия или калия,...
-
Получение, Промывка - Особенности получения и применения золота
Для получения золота используются его основные физические и химические свойства: присутствие в природе в самородном состоянии, способность реагировать...
-
Физико-химические свойства ртути - Ртуть: свойства и токсичность
Ртуть (Hg) - химический элемент II группы периодической системы элементов Д. И. Менделеева; атомный номер 80, относительная атомная масса 200,59; в...
-
Обзор литературы, Роль лизина в питании человека и животных - Этапы получения лизина
Роль лизина в питании человека и животных Лизин (б, е-диаминокапроновая кислота) - незаменимая основная алифатическая аминокислота. Это специфичная...
-
Строение и физико-химические свойства аскорбиновой кислоты - Аскорбиновая кислота
Аскорбиновая кислота (Acidumascorbinicum) - это белые кристаллы с резким кислым вкусом. Молекулярная масса = 176,13. Температура плавления аскорбиновой...
-
Синтез в живых системах - Простагландины. Строение. Биороль
Простагландины не накапливаются в тканях, а синтезируются в ответ на биологический стимул из полиненасыщенных жирных кислот: простагландины с одной...
-
Химическая связь - Квантовые концепции в химии
Химия изучает процессы превращения молекул при воздействиях и при воздействии на них внешних факторов (теплоты, света, электрического тока, магнитного...
-
Введение, Общие сведения о фосфорной кислоте - Общие сведения о фосфорной кислоте
Фосфорная кислота является основным сырьем для производства фосфорных удобрений, кормовых добавок, инсектицидов и других фосфорсодержащих продуктов....
-
Факторы, влияющие на состав и свойства молока - Химия и физика молока
1. Порода коров 2. Стадия лактации 3. Здоровье коров 4. Режим кормления 5. Другие факторы. Выход и качество молочных продуктов, определяемые составом...
-
Физико-химические свойства хрома - Хром. Элемент периодической системы химических элементов
Хром является серебристо-белым, твердым, блестящим, но в то же время довольно хрупким металлом. Ранее считалось, что хром практически не обладает...
-
Химическая связь - это взаимное сцепление атомов в молекуле и кристаллической решетке в результате действия между атомами электрических сил притяжения....
-
Рассмотрим химические свойства тиосульфата натрия. Изучим его окислительно-восстановительные свойства, взаимодействие с кислотами, комплексообразование....
-
История открытия тиосульфата натрия, Строение молекулы и ее влияние на свойства - Тиосульфат натрия
Имя и время открытия тиосульфата натрия нам не известны. Во всяком случае в истории химии об этом не упоминается. Но в конце XVII и в начале XVIII века,...
Особенности переваривания и всасывании жиров у детей - Обмен и функции липидов