Водоросли: общая характеристика, происхождение, систематика, распространение, хозяйственное значение - Флора, растительность, фитоценоз
В составе полцарства выделяют до 10 и больше отделов водорослей; сюда же включают и отдел лишайники.
Основные признаки следующие: 1) клетки крупные, 10--100 мкм; 2) ядра нормальной структуры, с ядрышком и мембраной; 3) имеются хондрносомы и пластиды, в ходе фотосинтеза освобождается молекулярный кислород, атмосферный азот не фиксируется; 4) как правило, клетки делятся посредством митоза. Имеется также половой процесс, а следовательно, уменьшение вдвое числа хромосом в ходе мейоза.
Древнейшие представители растительного мира, возникли около 1,5 млрд. лет назад. Общее число видов водорослей -- около 35 тыс. Благодаря постоянству условий жизни в водной среде, в которой водоросли возникли и пережили целые геологические эпохи, они сохранились до наших дней в формах, мало отличающихся от первоначальных. В процессе филогенеза организмы эволюционировали от одноклеточных и колониальных к мощным, сложно организованным гигантам, достигающим десятков метров длины. У высокоорганизованных представителей можно наблюдать начало некоторой внутренней дифференциации тела на ткани: покровную, ассимиляционную, запасающую, проводящую, механическую. Таким образом, у водорослей -- два контрастных полюса организации: па одном они представлены примитивными одноклеточными и колониальными формами и близки к простейшим животным, на другом -- многоклеточными, иногда гигантскими формами.
Большинство водорослей содержит хлорофилл и питается автотрофно, но нередко зеленая окраска их замаскирована другими пигментами. Небольшое число водорослей характеризуется как сапрофиты и паразиты. Тело водорослей, как и других низших растений, именуют талломом, поскольку оно не дифференцировано на настоящие органы и ткани. В большинстве случаев вегетативные клетки таллома снаружи покрыты твердой стенкой, состоящей из целлюлозы и пектиновых веществ. Нередко наружная поверхность клеточной стенки одета слизью или инкрустирована кремнеземом. У более примитивных одноклеточных или пальмеллоидных водорослей (например, у пирофитовых) клетки голые. Цитоплазма заполняет всю полость клетки или расположена постенно. Одна крупная или несколько мелких вакуолей заполнены клеточным соком. В клетке находится одно или несколько ядер и хроматофоры (пластиды), содержащие пигменты. По форме хроматофоры необычайно разнообразны (пластинчатые, спиральные, лентовидные, чашевидные, сетчатые, зернистые, звездчатые и т. д.), что служит важным признаком при определении водорослей. В хроматофорах у некоторых расположены белковые тельца -- пиреноиды, вокруг которых откладывается запасной продукт в виде крахмала или близкого к нему углевода. У эвгленовых таковым является парамилон. Кроме крахмала, в качестве запасного продукта может образоваться масло. У синезеленых водорослей оформленного ядра нет, как нет хроматофоров и обычных для других водорослей органелл. Поэтому синезеленые рассматриваются в предыдущем подцарстве.
Для водорослей характерны все типы размножения: вегетативное, бесполое и половое. Вегетативное происходит путем деления одноклеточных организмов, распадом колоний -- у колониальных, частями таллома -- у многоклеточных, а иногда путем образования специальных органов вегетативного размножения (например, клубеньков у харовых). Настоящее бесполое размножение осуществляется с помощью зооспор или спор-- одноклеточных образований, возникающих внутри вегетативных клеток или в особых органах--зооспорангиях (или спорангиях) путем деления содержимого. Зооспоры в отличие от спор имеют двигательные органы -- жгутики. Вскоре после появления зооспоры сбрасывают жгутики, покрываются клеточной стенкой и прорастают в новую особь. Как правило, бесполым способом водоросли размножаются в условиях, благоприятных для роста.
По мере ухудшения условий существования (высыхание, низкая температура) они приступают к половому размножению, которое происходит путем слияния двух, в большинстве случаев специализированных половых клеток -- гамет -- в одну клетку -- зиготу. В зиготе протопласты сливаются, накапливаются запасные продукты, а на поверхности развивается довольно толстая клеточная стенка. Затем зигота опускается на дно водоема, где она способна переносить неблагоприятные условия в состоянии покоя. В некоторых же случаях сформировавшаяся зигота сразу прорастает. При прорастании зиготы возможны два варианта: 1) зигота прорастает в новую особь; 2) в зиготе образуются зооспоры, которые прорастают по выходе в окружающую среду.
Половое размножение возможно в различных формах. При изогамии обе гаметы однаковы как по внешнему виду, так и по величине (изогаметы), но различаются физиологически (гетероталличные гаметы). При гетерогамии гаметы различаются не только гетероталлизмом, но и величиной, при этом женская гамета крупнее мужской. Изо - и гетерогаметы образуются путем редукционного деления содержимого обычной вегетативной клетки.
При наиболее совершенном у водорослей способе полового размножения -- оогамии -- женская гамета (яйцеклетка) представляет собой крупную неподвижную клетку с большим количеством запасных продуктов; мужская гамета -- сперматозоид -- маленькая клетка двух типов: амебоидного или монадного (жгутикового).
Половые клетки у водорослей развиваются в специальных, как правило, одноклеточных органах полового размножения (гаметангиях): яйцеклетки -- в оогониях, сперматозоиды -- в антеридиях. Яйцеклеток образуется одна или несколько, сперматозоидов -- много. Нередко у водорослей наблюдается половой процесс в виде конъюгации (зигогамии)-- слияния протопластов двух вегетативных клеток гетсроталличных особей (спирогира -- Spirogyra и др.).
У более примитивных групп (пирофитовые и др.) встречается гологамия и мерогамия. Однако половой процесс не всегда ведет к численному увеличению особей. Поэтому приходится говорить о половом воспроизведении. Конечный результат любого способа полового размножения (воспроизведения) -- слияние протопластов и образование зиготы. При этом удваивается число хромосом, т. е. восстанавливается диплоидная (2п хромосом) фаза. В типичном случае из зиготы развивается бесполая диплоидная фаза -- спорофит, на котором формируются органы бесполого размножения -- зооспорангии. В результате редукционного деления содержимого зооспорангия образуются гаплоидные (п хромосом) зооспоры, прорастающие в гаметофит, на котором образуются органы полового размножения -- гаметангии, продуцирующие гаметы. Гаметофит может быть как обоеполым, так и раздельнополым. Таким образом, у водорослей можно проследить 3 типа хорошо выраженной смены фаз развития: половой и бесполой.
1. Гаплоидный тип. Вегетативное тело всегда гаплоидно; диплоидна лишь зигота (первое деление ее ядра -- редукционное). Этот тип развития широко распространен среди водорослей (вольвокс, улотрикс и др.). Он чаще всего представлен следующей схемой:

В качестве примера гаплоидного типа развития рассмотрим зеленую нитчатую водоросль улотрикс. Индивидуумы этого вида состоят из гаплоидных клеток. При благоприятных условиях они быстро размножаются бесполым путем посредством зооспор. Осенью отдельные вегетативные клетки нити превращаются в гаметангии, внутри которых развиваются двужгутиковые изогаметы (гологамия). Выйдя из гаметангиев и проплавов некоторое время свободно, гаметы сливаются, образуя четырехжгутиковую зиготу. После сбрасывания жгутиков и развития толстой клеточной стенки зигота впадает в состояние покоя. Весной первое деление зиготы редукционное, в результате чего образуются 4 клетки, каждая из которых прорастает в новую особь. Таким образом, у улотрикса диплоидна только зигота. Весь жизненный цикл растение проходит в гаплоидной фазе. Приведенная схема гаплоидного развития достаточно типична для зеленых водорослей, но встречается и среди представителей других отделов.
2. Диплоидный тип. Вегетативное тело растения всегда диплоидно, поскольку редукционное деление свершается только при образовании гамет. Возникающая зигота сразу же (без периода покоя!) прорастает в новую вегетативную особь. Пример этого типа развития -- сифоновые водоросли, например бриопсис и кодиум.
Рассмотрим кодиум. Эта водоросль произрастает в Черном море на глубине до 10 м. Особи располагаются группами, формируя заросли. Их шнуровидное тело достигает 40--50 см длины. Ветвление дихотомическое. В период полового размножения на женских экземплярах вырастают гаметангии с подвижными двужгутиковыми мегагаметами, а на мужских экземплярах -- гаметангии с микрогаметами. Оплодотворение происходит в воде, после чего формируется зигота. При прорастании зиготы происходит митотическое деление, и из нее развивается также диплоидное растение. Редукционное деление наблюдается лишь в гаметангиях перед образованием гамет. Таким образом, весь жизненный цикл Codium проходит в диплоидной фазе.
3. Гаплоидно-диплоидный тип. Характерно чередование фаз развития ("поколений"). Редукционное деление совершается при образовании спор на диплоиде -- спорофите (спорофаза -- 2п хромосом). Из споры развивается гаплоид -- гаметофит (гаметофаза -- п хромосом), на котором образуются гаметы. Из зиготы (2п хромосом) без периода покоя развивается спорофит. Хорошие примеры этого типа развития -- энтероморфа -- Enteromorpha и ульва -- Viva (зеленые водоросли, порядок Улотриксовые). Ульва имеет форму двухслойной пластины с гофрированными краями. После слияния гамет из зиготы (2п хромосом) в результате митотического деления развивается диплоидный же спорофит. На спорофите формируются зооспорангии, где после редукционного деления появляются моноплоидные зооспоры.
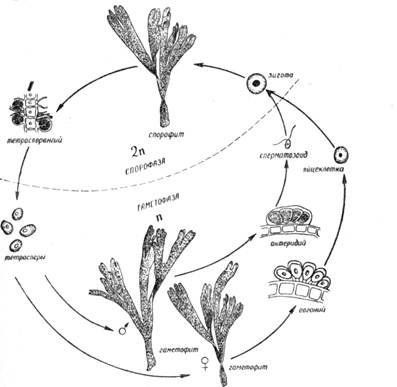
Рис. 1. Схема изоморфной смены фаз развития диктиоты
Далее из зооспор развиваются гаплоидные раздельнополые гаметофиты. Раздельнополость в данном случае выражается гетероталличностью (раздельнополостью талломов), поскольку гаметы в пределах одной особи не копулируют. Таким образом, совершается последовательная смена фаз развития -- диплоидной спорофазы и гаплоидной гаметофазы. При этом важно отметить, что в данном случае спорофит (2п) и гаметофит (п) внешне не различаются. В тех случаях, когда и гаметофит-гаплоид и спорофит-диплоид морфологически неразличимы, говорят об изоморфной смене фаз развития, которая наблюдается у некоторых бурых водорослей, например у диктиоты (рис. 2).
Это крупная лентовидная морская водоросль с дихотомическйм ветвлением. На спорофите из поверхностных клеток развиваются крупные тетраспорангии. Тетраспоры (не зооспоры) голые, неподвижные, образуются в результате редукционного деления. С помощью тетраспор совершается бесполое размножение диктиоты. Попав в воду, тетраспора прорастает в гаплоидное растение (гаметофит).
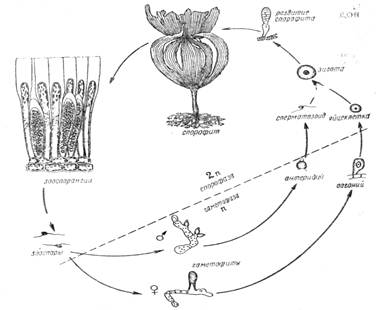
Рис. 2. Схема гетероморфной смены фаз развития ламинарии
На этих гаметофитах развиваются только органы полового размножения: на мужских экземплярах-- антеридии с многочисленными одножгутиковыми сперматозоидами, на женских -- оогонии с одной крупной яйцеклеткой в каждом. Спорангии на этих особях никогда не образуются. Яйцеклетка оплодотворяется в воде. Из зиготы вырастает диплоидное растение -- спорофит, служащее для бесполого размножения тетраспорами, но морфологически не отличающееся от гаметофита. Таким образом, у диктиоты хорошо выражена смена фаз развития: бесполая (спорофаза) и половая (гаметофаза), хотя растения при этом морфологически одинаковы.
Иное соотношение фаз развития наблюдается у бурой водоросли ламинарии. Виды этого рода широко распространены, часто встречаются в северных морях. Вегетативное тело представляет собой таллом в виде крупной листовидной или пальчаторассеченной пластинки и стеблевидного черешка. На базальной части "черешка" развиваются ризоиды, которыми растение прикрепляется к субстрату. В отдельных случаях ламинарии достигают 6--8 м высоты, а иногда и больше. На гигантском талломе развиваются зооспорангии -- органы бесполого размножения. Этот таллом представляет собой спорофазу (спорофит, точнее зооспорофит). Освободившиеся зооспоры прикрепляются к камням и прорастают в нитевидные микроскопические заростки. Это гаметофаза (л хромосом), причем гаметофиты здесь раздельнополые. У ламинарии спорофит и гаметофит по морфологии и размерам резко различаются (гетероморфия).
После длительного периода развития на женском гаметофите вырастают оогонии с одной яйцеклеткой внутри, а на мужском -- антеридии с многочисленными двужгутиковыми сперматозоидами. После оплодотворения, которое происходит в воде, зигота быстро развивается в крупное растение - диплоидный спорофит (2л хромосом).
Происхождение водорослей. Вопрос о происхождении, ходе развития водорослей и родственных связях между ними все еще не решен. Наиболее популярны две теории: 1) происхождение водорослей от одноклеточных автотрофных жгутиковых -- Flagellatae, близко стоящих к монадным водорослям, и 2) симбиотнческое происхождение: хлоропласта -- предъядерные водоросли, митохондрии -- аэробные бактерии.
При выяснении родственных взаимосвязей между отделами водорослей и уровня их исторического развития принимают во внимание следующие показатели: строение клетки, способы размножения, физиолого-биохимические особенности.
Наиболее высокоразвитые водоросли -- харовыс (многоклеточные органы полового размножения).
Последовательность изложения отделов водорослей (с. 96) мы даем, исходя из теоретических основ системы А. Пашера и Б. Фотта. Современные альгологи считают возможным видоизменять их систему в соответствии с новыми данными, которые дают биохимия и электронная микроскопия.
Большинство водорослей произрастает в воде, преимущественно морской. Они плотной массой как бы опоясывают побережья морей и океанов. Одни из них свободно плавают, составляя существенную часть взвешенных в толще воды организмов -- планктона (средняя масса особи планктонных видов 0,000006 мг). Водоросли, свободно лежащие на дне или прикрепляющиеся к нему, составляют часть донных организмов -- бентоса.
Поскольку водоросли -- фотосинтезирующие растения, то одно из основных условий их существования -- свет. В морях массовое распространение водорослей наблюдается на глубине до 30 м. Однако наиболее теневыносливые бурые и красные водоросли достигают глубины 100--200 м, а отдельные виды -- 500 м. Известны и более глубоководные виды. Некоторые водоросли сохраняют жизнеспособность при очень низких, а другие при высоких температурах. Так, в полярных и высокогорных странах они живут на снегу, нередко окрашивая его в красный, зеленый, бурый, желтый цвета (снежная хламидомонада). Многие водоросли не погибают под покровом снега или льда.
Отдельные виды, попадая вместе с бактериями на бесплодные субстраты, являются пионерами их заселения. Водоросли живут па почве, в почве и даже в атмосферном воздухе, например некоторые виды хлореллы. Многие почвенные водоросли активно участвуют в процессе почвообразования. Так, при подзолообразовании в почве накапливается значительное количество зеленых (улотриксовых) водорослей, при дерновом процессе -- диатомовых, при болотном -- зеленых, на глинистых почвах пустынь -- синезеленых и т. д. Азотфиксирующие водоросли накапливают в почве азот.
Водоросли не только обогащают воду кислородом, но служат превосходным кормом для рыб и водоплавающих птиц. Высока питательная ценность водорослей, особенно планктона (до 30 % белка и до 60 % углеводов, много витаминов, жиров и ценных минеральных солей). Муку из бурых водорослей используют на корм сельскохозяйственным животным. В северных приморских районах и на Дальнем Востоке на корм идут ламинарии, фукус и другие виды в свежем и силосованном виде. Их поедают лошади, крупный рогатый скот, свиньи. Тонна муки из сухих водорослей содержит около 180 кг солей калия, 16 кг органического азота, 10 кг фосфора и другие вещества, необходимые для развития животных. Водоросли применяют как быстродействующие удобрения в свежем и компостированном виде. Зола бурых и красных водорослей -- сырье для получения брома и иода, сами водоросли используются для получения агара, употребляемого в кондитерской промышленности и для изготовления питательных сред в микробиологии.
Морские водоросли нередко используют в текстильной и бумажной промышленности.
Похожие статьи
-
Основные классы красных водорослей, Бангиевые. Порфира - Красные водоросли и их характеристика
Бангиевые. Порфира Класс бангиевых объединяет одноклеточные, колониальные и многоклеточные формы паренхимного строения. Рост у них диффузный, в...
-
Строение красных водорослей - Красные водоросли и их характеристика
Споры и гаметы багрянок лишены жгутиков, а цикл их развития включает не две, как у других водорослей, а три стадии. После слияния гамет из зиготы...
-
Разновидности канальцев яичка и придатка представлены: I. Сперматогенные, или извитые семенные, канальцы охарактеризованы в таблице 1.1....
-
Простейшие с организацией жгутиконосцев - Строение, размножение и жизненные циклы простейших
Данная группа включает более 10 типов. Свободноживущие и паразитические животные. Одиночные, реже колониальные. Размеры животных и форма тела...
-
Характеристика тип Апикомплексы - Апикомплексы
Это большая группа паразитических простейших, насчитывающая около 4800 видов. Среди них много опасных паразитов человека и животных. К апикомплекс...
-
Классификация типов дробления: - Дробление, его характеристика у разных животных. Типы бластул
Существует Несколько типов классификации процесса дробления. I . По характеру образования и расположению бластомеров: 1. Полное ( Голобластическое ) -...
-
Общая характеристика В последующем половые шнуры срастаются с канальцами I почек. Из половых шнуров образуются эпителиосперматогенный слой извитых...
-
Плазма крови - это раствор, состоящий из воды (90-92%) и сухой остаток (10 - 8%), состоящий из органических и неорганических веществ. (рис. 1)...
-
Основные свойства фитоценозов В данном разделе рассматриваются только естественные фитоценозы и при этом коренные, или климаксовые (объяснение этого...
-
Форма этих организмов удлиненно-веретенообразная или нитевидная. Кожно-мускульный мешок состоит из многослойной, плотной, эластичной и нерастяжимой...
-
Бластула - Дробление, его характеристика у разных животных. Типы бластул
У многих яйцеклеток еще на ранних стадиях дробления внутренние концы бластомеров расходятся, и между ними возникает сначала небольшая, а затем все...
-
Типы бластул - Дробление, его характеристика у разных животных. Типы бластул
Имеется 5 основных типов бластул: - Целобластула. - Бластоциста. - Амфибластула. - Дискобластула. - Перибластула. Целобластула характерна для иглокожих и...
-
Механизм дробления - Дробление, его характеристика у разных животных. Типы бластул
Клеточный цикл соматических клеток делится на четыре стадии (фазы). За митозом (М) следует пресинтетический период (G1), после которого происходит синтез...
-
Обычно гликозиды классифицируют по типу агликона. Основные классы гликозидов перечислены ниже. Тиольные гликозиды (тиоцианатные, изотиоцианатные, сульфо...
-
"Наша способность к самообмену по поводу работы собственного мозга почти безгранична главным образом потому, что часть, о которой мы можем сообщить,...
-
Общая характеристика растущего организма С биологических позиций, период роста организма можно разделить на три этапа: - детский - с рождения до 11-12...
-
Психические изменения при эндокринных заболеваниях носят полиморфный характер, но их развитие подчиняется определенным закономерностям, которые сводятся...
-
Строение глаз кошки - Общая характеристика кошек
Строение глаза у кошки в основном такое же, как у человека. Зрение основано на способности организмов обнаруживать свет видимую часть электромагнитного...
-
Общая характеристика споровиков. Приспособления к паразитизму - Апикомплексы
Класс споровиков представляет собой обширную группу простейших организмов, ведущих исключительно паразитический образ жизни. Приспособление к паразитизму...
-
Глобализация - процесс слияния национальных экономик в единую, общемировую систему, основанную на быстром перемещении капитала, новой информационной...
-
Наиболее важной буферной системой плазмы является бикарбонатный буфер, состоящий из слабой угольной кислоты (рК1 6,1) и ее кислого аниона бикарбоната....
-
Формирование человека происходило под определяющим воздействием социальных факторов. Поэтому как объект естественнонаучного исследования он представляет...
-
Питание и упитанность. Лещ является типичным бентофагом, однако наряду с хирономидами, бокоплавами, моллюсками и другими бентосными организмами лещ...
-
Хозяйственное значение - Биологическая характеристика черноморской хамсы
Черноморский анчоус - одна из важных промысловых рыб Черного и Азовского морей, составлявшая в довоенные годы около четверти общего улова(рисунок 5). И...
-
Развитие половой системы - длительный процесс, который не завершается в эмбриональном периоде, а продолжается после рождения, вплоть до достижения...
-
Личинка - Разнообразие личинок у разных организмов
Эмбриональным периодом принято называть отрезок времени от начала развития до рождения или до момента выхода зародыша из яйцевых оболочек. В конце...
-
Заключение - Тканевая инженерия в медицине
Клеточная (тканевая) инженерия -- отрасль биотехнологии, в которой используют методы выделения клеток из организма, трансформации их и выращивания на...
-
Согласно одному из распостраненных определений, гестоз представляет собой синдром полиорганной недостаточности, возникающей при беременности, в основе...
-
Этап. Филогенетическая (эволюционная) систематика - Систематика растений
Классификация основывается не только на общих морфологических или анатомических признаках растений, но и учитывает особенности и общность происхождения...
-
Апоптоз - Характеристика основ иммунологии
Пролиферация клеток Количество клеток в ткани регулируется двумя процессами-- Пролиферацией Клеток и " Программированной, или физиологической, гибелью...
-
БАЛАНТИДИЙ - Balantidium coli - возбудитель балантидиаза - антропозооноза. Географическое распространение - повсеместно. Локализация - толстый кишечник,...
-
Образовательные ткани (меристемы) - Характеристика отдельных видов растительных тканей
Важный признак, отличающий растения от животных, - рост в течение всей жизни. Рост растений ограничивается сезонными изменениями, а не возрастом. Этот...
-
Взаимопревращения пластид - Пластиды: строение, происхождение, взаимопревращения
В эволюционном смысле первичным, исходным типом пластид являются хлоропласты, из которых при расчленении тела растений на органы произошли пластиды...
-
Введение - Дробление, его характеристика у разных животных. Типы бластул
Оплодотворение играет чрезвычайно важную роль в развитии, но оно лишь только первая его ступень. Зигота с ее новым генетическим потенциалом и новым...
-
Жизненный цикл клетки Жизненный цикл-- это время существования клетки от момента ее образования путем деления материнской клетки до собственного деления...
-
Эхинококк Echinococcus granulosus - возбудитель эхшюкоккоза. Половозрелая форма имеет головку с крючьями и 3-4 членика разной степени зрелости. Последний...
-
Жук биология вредитель экосистема Усачим, или дровосемки (лат. Cerambycidae) -- разветвленное и многочисленное (пятое по количеству видов) семейство...
-
Жизненный цикл клетки отражает все закономерные структурно-функциональные изменения, происходящие с клеткой во времени. Жизненный цикл - это время...
-
Заболевания, относящиеся к дифференцированной дисплазии соединительной ткани, встречаются в популяции достаточно редко. Например, несовершенный...
-
Мухи - известны как наиболее активные механические переносчики возбудителей заболеваний. Как и у всех двукрылых, у мух одна пара передних крыльев;...
Водоросли: общая характеристика, происхождение, систематика, распространение, хозяйственное значение - Флора, растительность, фитоценоз