Этап. Филогенетическая (эволюционная) систематика - Систематика растений
Классификация основывается не только на общих морфологических или анатомических признаках растений, но и учитывает особенности и общность происхождения растительных видов.
С развитием морфологии растений искусственная систематика растений уступила свое место "естественной", основанной на совокупности признаков. Первая "естественная система" была создана в 1789 г. Однако "естественная систематика" не была еще естественной в современном смысле, так как она не была еще эволюционной[7]. Авторы естественных систем продолжали верить в постоянство видов. В естественных системах растения объединялись на основании "сродства", или "родства", под которым понималось, однако, не родство по происхождению, а лишь внешнее и часто поверхностное сходство.
В естественных системах соединяются такие растения, которые обнаруживают наибольшее внешнее сходство между собой. В результате естественная систематика часто объединяла аналогичные эволюционные стадии или сходные верхушки разных филогенетических ветвей, т. е. она строила свои рубежи поперек течения эволюции. Тем не менее, многие построения естественной систематики предвосхитили выводы эволюционной систематики.
После торжества эволюционной идеи в биологии естественная систематика стала постепенно уступать свое место эволюционной, или филогенетической, систематике. Начался новый этап в ее развитии. Употреблявшийся и ранее термин "родство" получил новое значение, и перед систематикой возникли новые цели. Основной задачей систематики является теперь построение такой системы классификации, которая отражала бы родственные, т. е. эволюционные, взаимоотношения между организмами.
Современная систематика развивается в тесной связи с другими биологическими науками и широко пользуется как их фактическим материалом и идеями, так и методами исследования, в том числе экспериментальными.
Наиболее ранние и очень отрывочные ископаемые остатки покрытосеменных (пыльца, древесина) известны из юрского геологического периода. Из нижнемеловых отложений известны тоже немногочисленные достоверные остатки покрытосеменных, а в отложениях середины мелового периода они встречаются сразу в больших количествах и в значительном разнообразии форм, которые все принадлежат ко многим различным ныне живущим семействам и даже родам.
В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений - кейтониевые, семенные папоротники, беннеттиты, гнетовые.
У кейтониевых были завязь, рыльце, но у них завязь формировалась иначе, чем у покрытосеменных; у них не было даже подобия цветков, спорофиллы их простые и, вероятно, они представляют слепую ветвь эволюции. У беннеттитов были обоеполые своеобразные "цветки", но не было пестиков, и семена их были лишь скрыты между бесплодными чешуями, а не находились внутри плодов, образованных мегаспорофиллами. У семенных папоротников не было цветков, не было покрытосемянности.
Теория происхождения покрытосеменных из гнетовых предполагает, что наиболее примитвные покрытосеменные имели мелкие однополые цветки без околоцветника или с невзрачным околоцветником. Но по ряду соображений в настоящее время более примитивными цветками считают крупные, обоеполые цветки.
Поэтому можно предположить, что предками современных покрытосеменных были какие-то вымершие, очень примитивные голосеменные с обоеполыми цветками типа шишки (стробилами), в которых на длинном цветоложе (оси) были спирально расположены свободные (несросшиеся друг с другом) листочки однородного околоцветника, микроспорофиллы (тычинки) и мегоспорофиллы (плодолистики). В системе голосеменных эта группа должна была стоять где-то между семенными папоротниками и уже более специализированными беннеттитами и саговниками.
Покрытосемянность несомненно представляла большое преимущество в смысле защиты семяпочек и развивающихся семян от всяких неблагоприятных внешних воздействий и в первую очередь от сухости воздуха. Но одной покрытосемянностью все же трудно объяснить быстрое мощное развитие покрытосеменных и вытеснение ими господствовавших ранее на земле архегониальных растений.
Русский ботаник М. И. Голенкин высказал (в 1927 г.) интересную гиппотезу о причинах победы покрытосеменных в борьбе за существование. Он предполагает, что в середине мелового периода по каким-то общим космогоническим причинам по всей Земле произошло резкое изменение освещения и влажности воздуха. Густые облака, окутовавшие ранее постоянно Землю, рассеялись и дали доступ ярким солнечным лучам, в связи с чем резко увеличилась сухость воздуха. Громадное большинство высших архегониальных растений того времени, не приспособленных и не сумевших приспособиться к яркому освещению и сухости воздуха, начало вымирать или резко сократило области своего распространения (кроме хвойных, наиболее ксерофитных).
Наоборот, покрытосеменные, имевшие до того очень ограниченное распространение и представление небольшим числом форм, выработали способность хорошо переносить яркий солнечный свет и сухость воздуха. Это обстоятельство, а также чрезвычайная эволюционная пластичность их, способность к возникновению разнообразнейших приспособлений к различным внешним условиям и обусловили быстрое победное распространение покрытосеменных по всей Земле и вытеснение ранее господствовавших групп высших архегониальных растений.
Цветковые, или покрытосеменные - сравнительно молодая группа высших растений, появившаяся в меловом периоде, стремительно размножившаяся в середине - конце мела и с тех пор занимающая господствующее положение на суше. Проблеме происхождения цветковых посвящено огромное количество публикаций, однако в этом вопросе по-прежнему много неясного. Общепризнано, что цветковые произошли от голосеменных.
Д. Д. Соколов и А. К. Тимонин подробно рассмотрели основные теории происхождения покрытосеменных. Авторы обращают внимание на парадоксальный факт: цветы покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Поэтому и сформулировать, что такое цветок, весьма трудно. Тем не менее, ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных.
Впрочем, если не рассматривать мужские цветки, у остальных все-таки есть общая черта, отличающая цветок покрытосеменных от фруктификаций голосеменных.
У цветковых плодолистики свернуты и образуют, во-первых, рыльце, улавливающее пыльцу, во вторых - замкнутую полость (завязь), в которой развиваются семязачатки.
У некоторых голосеменных есть нечто похожее - замкнутые капсулы с семязачатками, однако у них нет завязи (пыльцу улавливают сами семязачатки при помощи длинных отростков, высовывающихся из узкого отверстия капсулы).
К тому же исходный листочек, несущий семязачатки (мегаспорофилл, у цветковых ему соответствует плодолистик) свернут у голосеменных нижней стороной внутрь, а у цветковых - наоборот, верхней стороной внутрь. Соответственно и семязачатки сидят на разных сторонах листочка.
Эти различия представляются настолько радикальными, что большинство ботаников не видят возможности постепенного перехода из одного состояния в другое и предполагают сальтационное превращение (одним скачком).
Это вполне возможно, если учесть, что развитие фруктификаций у растений управляется небольшим числом генов-регуляторов, мутации в которых действительно могут приводить к скачкообразным изменениям плана строения. Аналогичная ситуация наблюдается и у животных, план строения которых в основном определяется небольшим набором генов-регуляторов.
В начале XX века была популярна теория происхождения цветковых от беннеттитов - вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) - в отличие от других голосеменных, имеющих однополые фруктификации.
Из стробилов беннеттитов, действительно напоминающих цветы, ботаники и пытались вывести типичный для покрытосеменных обоеполый цветок. Но тут возникла проблема: у беннеттитов не обнаружилось никаких структур, из которых можно было бы вывести плодолистики. У них вообще нет мегаспорофиллов, а семязачатки сидят прямо на оси стробила. Проблема казалась неразрешимой.

Рисунок 1. Лист беннеттитового Pterophyllum (поздний триас)
В 80-х годах XX века крупнейший российский палеоботаник С. В. Мейен выдвинул гипотезу, разрешающую это противоречие. Он предположил, что у каких-то беннеттитовых сальтационным путем произошла перестройка программы развития фруктификации. В результате ключевые элементы программы развития мужских элементов стробилы (микроспорофиллов, у цветковых это тычинки) стали реализовываться при развитии женских элементов.
Это привело к появлению химерных образований - микроспорофиллов, несущих семязачатки вместо пыльников. Именно из таких "мутантных" микроспорофиллов и образовались плодолистики. У беннеттитовых микроспорофиллы несут пыльники как раз на своей верхней стороне, в отличие от других голосеменных. Поэтому такая мутация сразу привела к появлению "правильных" прототипов для формирования плодолистиков, с семязачатками на нужной стороне.
Ничего невероятного в таком сценарии нет, исходя из того, что известно об общих принципах генетической регуляции развития. Гипотеза Мейена объясняет противоречие, казавшееся неразрешимым. Ее нельзя проверить классическими методами палеоботаники и морфологии (поскольку никаких переходных форм гипотеза Мейена не предполагает). Однако ее можно проверить методами сравнительно-генетического анализа.
Генетическая регуляция развития фруктификаций у семенных растений стала активно изучаться в начале 90-х годов XX века. Оказалось, что у всех цветковых главным регулятором развития цветка является ген LFY.
У голосеменных есть аналогичный ген (Leaf family), который экпрессируется (работает) на ранних стадиях развития мужских фруктификаций. Кроме того, у голосеменных есть еще один сходный ген (Needle family), отсутствующий у цветковых, который участвует в формировании женских фруктификаций. У большинства голосеменных, напомним, фруктификации раздельнополы. При появлении цветковых, таким образом, "женский ген" был утрачен, а "мужской" стал руководить развитием всего цветка, с его мужскими и женскими частями.
На этом основании Фролик и Паркер (Frolich, Parker, 2000) выдвинули "преимущественно мужскую теорию" происхождения цветка. Они предположили, что в результате утраты гена из семейства Needle у предков цветковых женские фруктификации перестали формироваться, а семязачатки стали развиваться на верхней стороне плоских микроспорофиллов. Казалось бы, это в общем то же самое, что предполагал С. В. Мейен.
Авторы "преимущественно мужской теории", однако, подчеркивают, что Мейен предполагал перенос признаков мужских структур на женские, а они говорят о переносе органов с места на место (семязачатков на микроспорофиллы). Но это, конечно, больше похоже на игру в слова, а не на существенное различие двух теорий. Различие на самом деле в другом, а именно в вопросе о предковой группе цветковых.
Эволюционные реконструкции, основанные на морфологии, подтверждали родство покрытосеменных и беннеттитовых (а также родственных беннеттитам гнетовых, которые дожили до наших дней). Однако реконструкции, основанные на молекулярных данных, показали иное. Они показали, что все дожившие до современности голосеменные представляют собой монофилетичную группу (т. е. группу, происходящую от одного общего предка и включающую всех без исключения потомков этого предка).
Это значит, что последний общий предок всех современных голосеменных, в том числе гнетовых (а стало быть и беннеттитов тоже) не был предком покрытосеменных.
Таким образом, предка цветковых нужно искать среди других вымерших голосеменных, более древних, точнее, обособившихся раньше, чем беннеттитовые. Фролик и Паркер предложили свою кандидатуру - користоспермовые семенные папоротники (Corystospermataceae). У них, правда, микроспорангии (пыльники) располагались на нижней, а не на верхней стороне микроспорофиллов.
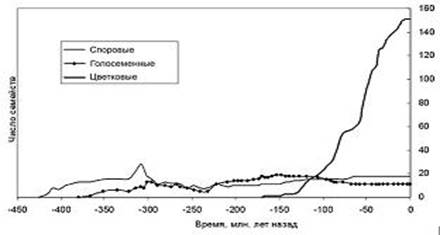
Рисунок 2. Динамика разнообразия основных групп наземных растений во времени
Д. Д. Соколов и А. К. Тимонин также приводят доводы в пользу того, что строение семязачатков користоспермовых не позволяет вывести из них семязачатки покрытосеменных. "Подходящие" семязачатки есть у другой группы голосеменных - у кейтониевых, но зато у них нет ничего похожего на плоские микроспорофиллы, из которых можно было бы вывести плодолистики.
Авторы предполагают, что причина всех этих противоречий, вероятно, кроется в недостаточной надежности эволюционных реконструкций, основанных на молекулярных данных. По их мнению, голосеменные растения плохо подходят для молекулярно-филогенетического анализа, поскольку большинство семейств голосеменных давно вымерло. Мы, таким образом, имеем недостаточно репрезентативную выборку таксонов, что, как хорошо известно, может приводить к ошибочным молекулярным филогениям. В конечном счете, по мнению авторов, на сегодняшний день нет оснований для безоговорочного отрицания идеи Мейена о происхождении цветковых от беннеттитовых. Эту идею вполне можно сохранить в качестве рабочей гипотезы.
По вопросу о месте первоначального возникновения покрытосеменных высказывались разные предположения. Одни считают, что они впервые появились на гипотетическом тропическом материке, расположенном между Америкой, Азией и Австралией и впоследствии погрузившемся в воды Тихого океана. Другие считают колыбелью их области современной арктической суши, третьи - горы субтропической и умеренно теплой зоны северного полушария. Большинство ботаников в настоящее время считают, что первичные покрытосеменные были древесными растениями, имевшими невысокие стволы, моноподиально разветвлявшиеся на немногочисленные толстые ветви.
Из них уже развились более крупные симподиально ветвящиеся деревья с многочисленными толстыми и тонкими ветвями. Из древесных же форм в разное время и разных филогенетических линиях развивались кустарники, полукустарники и травянистые формы, вначале многолетние, затем в различных родах в связи со специфическими условиями климата и местообитаний - двухлетники и однолетники.
Благодаря большой пластичности покрытосеменных, у них в процессе эволюции выработалось огромное разнообразие вегетативных органов, особенно в листьях, многочисленные метаморфозы, а также бесконечное разнообразие в цветках и плодах.
Сложность и разнообразие химического состава и физиологических реакций также очень характерно для них.
Эволюция цветка, на строении которого главным образом базируется систематика покрытосеменных, говоря в общем и схематическом виде шла у них от цветков с длинным цветоложем (типа шишки) от обоеполых, актиноморфных со спиральным расположением свободных (несросшихся) и не фиксированных в числе членов, с верхней завязью и многочисленными семяпочками - к цветкам циклическим, зигоморфным, раздельнополым, со строго фиксированным числом более или менее сросшихся членов на плоском цветоложе, с нижней одногнездною завязью и немногими или одной семяпочкой. Эта эволюция цветка покрытосеменных происходила в разных эволюционных рядах их независимо друг от друга.
Анатомо-морфологическая характеристика отдела покрытосеменных. Покрытосеменные - самый крупный тип растений, к которому относится более половины всех известных видов. Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро - и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище - завязь, в которой развиваются семяпочки (макро - и мегаспорангии). После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя).
Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплойдный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства - более или менее типичный цветок с околоцветником. Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиты, а сосуды встречаются крайне редко.
За небольшими исключениями, все покрытосеменные растения имеют корень, стебель и листья.
Корень (radix) развивается под землей и служит растению для укрепления в почве и всасывания из нее воды и растворенных в ней неорганических веществ, необходимых для его нормального существования. Нормальные корни каждого растения образуют его корневую систему.
Различают два основных типа корневой системы: стержневая корневая система (дуб, наперстянка, одуванчик и др.) и мочковатая или пучковатая корневая система (все луковичные, такие, как чеснок, лук и др., а также и большинство злаков, таких, как кукуруза, пшеница и др.).
У некоторых растений корни видоизменены: мясистые корни (морковь, свекла, редька и др.), корневые клубни (большая часть болгарских ятрышников), клубневые корни (георгина, таволга, калужница и др.). Во всех видоизмененных таким образом корнях откладываются запасные пищевые вещества.
Стебель (caulis и cauloma), когда он нормальный, развивается над поверхностью земли. По стеблю проводится всосавшаяся из почвы вода с растворенными в ней неорганическими веществами наверх к листьям и возвращается вниз в корни вода с растворенными в ней органическими обработанными веществами; кроме того, на стебле и на его разветвлениях расположены листья и цветки.
Помимо нормальных надземных стеблей, у некоторых растений имеются видоизмененные подземные стебли, какими являются, например, корневища (rhizoma - касатик, ландыш майский, чемерица и др.), луковица (bulbus - подснежник, лук, чеснок и др.) и клубень (tuber - картофель, земляная груша и др.). И в видоизмененных стеблях откладываются запасные питательные вещества.
В зависимости от стойкости нормальных надземных стеблей растения делятся на два основных типа: древесные и травянистые. К первому типу причисляют деревья (дуб, черешня, береза и др.), кустарники (шиповник, терн, лещина и др.) и полукустарники (ежевика, черника и брусника, лаванда и др.).
Ко второму типу относятся многолетние травянистые растения, из подземных многолетних частей которых каждый вегетационный период вырастают надземные стебли, а на них образуются цветки, плоды и семена; зимой, однако, вся надземная часть таких растений погибает (белладонна, большинство видов горечавки, морозник, зверобой, ландыш майский и др.).
Двухлетние травянистые растения, у которых в первый год образуется прикорневая розетка листьев, во время зимы сохраняют свою подземную часть и только на второй год у них развивается стебель с цветками, плодами и семенами, однако зимой второго года они погибают (некоторые виды коровяка, иногда наперстянка шерстистая, иногда пастушья сумка, а из культурных растений - капуста, морковь и др.).
У однолетних травянистых растений за один вегетационный период из семени развивается взрослое растение, которое после образования цветков, плодов и семян, зимой или даже раньше погибает (дурман, очанка, золототысячник, а из культурных растений - кукуруза, пшеница и др.). Некоторые из однолетних растений могут перезимовать, когда семена их прорастают осенью, но это обычно наблюдается мягкой зимой.
Стебли растений бывают разветвленными (деревья, кустарники и многие травянистые) и неразветвленными (подснежник, ландыш, пшеница и др.). Разветвление у разных растений различное.
Листная система (лист - folium) является фотосинтезирующим аппаратом растения, где главным образом образуются органические вещества. Существует очень большое разнообразие, в особенности в отношении внешнего строения (морфологии) листа, а именно: прикрепление листьев и расположение их по стеблю, наличие или отсутствие черешка, влагалища или прилистников, устройство пластинки листа -- простая или сложная пластинка, форма пластинки листа у простых листьев, форма края пластинки, жилкование, консистенция листьев и др.
Лист покрытосеменных растений состоит из черешка (стебелька листа), листовой пластинки (лопасти) и прилистников (парных придатков, расположенных по обеим сторонам основания черешка). Место, где черешок примыкает к стеблю, называется влагалищем листа. Угол, образованный листом (черешком листа) и вышерасположенным междоузлием стебля, называется пазухой листа. В пазухе листа может образоваться почка (которая в этом случае называется пазушной почкой), цветок (называется пазушным цветком), соцветие (называется пазушным соцветием).
Цветок (flos). Цветок обеспечивает половое размножение или воспроизводство растения. Он представляет собой укороченный стебель с ограниченным ростом; на этом стебле видоизмененные листья располагаются в определенном порядке, в котором можно установить известные закономерности. Такие видоизмененные листья называют частями цветка.
Существенными частями являются тычинки и пестик, а несущественными - чашечка и венчик или простой околоцветник (перигон), когда нет обособленных чашечки и венчика (тюльпан, ландыш, подснежник и др.). Когда один и тот же цветок имеет и тычинки, и пестик, его называют обоеполым (гермафродитным); когда у цветка только тычинки - это однополый, мужской (тычиночный) цветок, а когда налицо только пестик - это однополый, женский (пестичный) цветок.
Когда мужские и женские цветки расположены на одном и том же индивиде, растение однодомное (грецкий орех, лещина, тыква и др.), когда они расположены на двух различных индивидах - двудомное (большинство видов ивы, конопля, крапива двудомная и др.).
Части цветка в обоеполых цветках располагаются в пять кругов по цветоложу, а именно, снаружи чашечка, затем, идя внутрь, венчик, тычинки (в два круга) и пестик. При простом околоцветнике лепестки расположены чаще всего в два круга. Каждая тычинка имеет пыльник, в гнездах которого образуется пыльца (цветочная пыльца), и тычиночную нить (не всегда).
Пестик состоит из длинной расширенной части - завязи, столбика (иногда отсутствует) и одного рыльца или нескольких рылец. Пестик образуется от срастания одного или большего числа плодолистиков. Если плодолистиков два или больше и каждый из них закроется сам по себе, получается сборный или сложный пестик (малина, лютик, морозник и др.). При срастании двух или большего числа плодолистиков (вместе) может образоваться одногнездная или многогнездная завязь.
В завязи образуется одна (черешня, слива, малина, подсолнечник и др.) или больше (томат, белладонна, груша, тыква и др.) семяпочек, которые после оплодотворения и развития в них зародыша превращаются в семена. В зависимости, главным образом, от способа опыления, то есть от способа перенесения пыльцы из пыльников тычинок на рыльце пестика (насекомыми, ветром, водой, птицами и др.) цветки бывают различно устроены - существует поразительное приспособление к переносчикам пыльцы.
В отношении симметрии цветки бывают полисимметричными или актиноморфными (черешня, подснежник, дурман и др.), оносимметричными или зигоморфными (фиалка, бобовые, наперстянка и др.) и ассимметричными или неправильными (валериана, хна и др.). Махровые цветки, которые почти всегда бывают бесплодными, получаются путем превращения тычинок в лепестки.
Соцветия (inflorescentiae). Очень редко цветки располагаются по одному на стебле (подснежник, мак, тюльпан, пион и др.)- Когда несколько или много цветков, между которыми нет нормальных листьев, я только прилистники, расположены в непосредственной близости друг от друга, они образуют соцветие. Соцветия покрытосеменных растений очень разнообразны, причем нередко они бывают сложными и комбинированными. Их можно разделить на две основные группы: 1) рацемозные (кисти, бокоцветные), с моноподиальным разветвлением; 2) цимозные (верхоцветные), с симподиальным или ложным, вильчатым разветвлением.
Семя (semen) и плод (fructus). Материальной предпосылкой оплодотворения является опыление, о котором уже упоминалось выше, а оплодотворение состоит в том, что обе оплодотворяющие клетки (спермии), образующиеся при прорастании пылинки на рыльце (проходя через пыльцевую трубочку), проникают в зародышевый мешок (эмбриосак) семяпочки и сливаются - одна с яйцеклеткой, а другая с вторичным ядром. После многократного деления оплодотворенной клетки образуется зародыш, а из оплодотворенного вторичного ядра образуется вторичная питательная ткань (эндосперм).
После окончательного развития и оформления зародыша семяпочка разрастается и превращается в семя с собственной оболочкой (testa), а завязь, в которой скрыта семяпочка (или семяпочки), так же разрастается, увеличивается и превращается в плод, у которого тоже есть своя оболочка (pericarpium). Строение и размеры семян очень разнообразные (орхидеи, ива, мак, тыква, каштан конский, кокосовая пальма и др.). По своему строению и размерам плоды еще более разнообразны, "следствие чего созданы различные классификации плодов. Одной из наиболее естественных и приемлемых является классификация, созданная на основании консистенции плодов и строения оболочки плода, получающейся при разрастании плодолистика или плодолистиков (когда завязь образована от сращения вместе двух или нескольких плодолистиков). Существует два основных типа плодов: сухие, у которых весь околоплодник деревянистый, кожистый или пленчатый, и сочные или мясистые, у которых весь околоплодник или часть его (пласт или пласты) сочные или мясистые.
Похожие статьи
-
Развитие систематики, этап. "Искусственная" систематика - Систематика растений
Основная ботаническая дисциплина - систематика[1] растений - разделяет многообразие растительного мира на соподчиненные друг другу естественные группы -...
-
Этап. Морфологическая систематика - Систематика растений
На первое место вышли морфологические признаки растительных организмов. На этом этапе можно говорить о возникновении первой "естественной" системы...
-
Семейство: бромелиевые. - Экологические группы растений по отношению к свету, воде и теплу
Гузмания: растение образует розетку длинных цель-нокрайних изогнутых листьев. Верхние листья на цветоносе (прицветники) ярко-красные или оранжевые до 4...
-
Этапы микроклонального размножения растений - Биотехнология микроклонального размножения особей
Процесс клонального микроразмножения можно разделить на 4 этапа: 1. Выбор растения-донора, изолирование эксплантов и получение хорошо растущей стерильной...
-
Кольчатые черви (кольчецы, аннелиды), тип червей. Тело поделено внутренними перегородками на сегменты, которым соответствует наружная кольчатость. Длина...
-
Условия произрастания - Экологические группы растений по отношению к свету, воде и теплу
В большинстве случаев розы теплолюбивы, но есть виды, растущие в условиях сурового климата. Розы предпочитают яркое освещение и при значительном...
-
Естественный отбор - Сущность эволюционной теории Чарльза Дарвина
По Дарвину, эволюция видов в природе обусловливается факторами, аналогичными тем, которые определяют эволюцию культурных форм. Предпосылкой эволюции...
-
Повилика - другое хорошо известное и широко распространенное растение-паразит, у которого нет ни корней, ни зеленых листьев. Само название этого паразита...
-
Петров крест - Растения-паразиты
Растение-паразит Петров крест чешуйчатый издавна поражал воображение людей. Петров крест является родственником еще одного растения-паразита - заразихи....
-
Малярийные плазмодии Plasmodium (кл. Споровики) - возбудители малярии. Известны следующие виды малярийных плазмодиев, паразитирующие у человека: P. vivax...
-
Естествознание на современном этапе - Естествознание - единая наука о природе
(современное естествознание) Современная наука ведет свой отчет с 10-20-х гг. ХХ в. Для нее характерна ассоциация математических, гуманитарных и...
-
Синантропные и культурные растения - Синантропная флора Дальнего Засвияжья г. Ульяновска
С деятельностью человека связано появление синантропных растений (антропофитов). Появились синантропные растения непреднамеренно помимо его воли человека...
-
Мать-и-мачеха - Мир лекарственных растений
Мать-и-мачеха - первый весенний цветок, появляющийся по берегам рек, оврагам, канавам, по глинистым обрывам и железнодорожным насыпям и в местах...
-
Сразу же отметим, что изменить положение своей БД для ответа на изменение внешней среды в водах с замедленным течением и тем более - в стоячих водах...
-
Процесс саморегуляции в дубраве проявляется в том, что все разнообразное население леса существует совместно, не уничтожая полностью друг друга, а лишь...
-
Одна из особенностей граната -- "незасыпаемость". Если стебель и ветви растения занесет песок, то растение пускает новые придаточные корни. Растения как...
-
Появление в научной литературе интереса к глобальным проблемам было связано с тем, что глобальные исследования касались единого процесса, происходящего...
-
Введение - Экологические группы растений по отношению к свету, воде и теплу
Светолюбивые растения. К светолюбивым растениям (гелиофитам) относятся виды, которые произрастают на открытых, хорошо освещенных солнцем местах, и не...
-
Взаимоотношения растений и насекомых - Биогеоценотический покров земли
Влияние фитофагов: - Пантофаги - группа животных, которые питаются всем, т. е. всеядные (кузнечик); - Полифаги - питаются растениями разных семейств...
-
Этапы развития нервной системы человека - Особенности нервной системы у детей
1)Дорзальная индукция или Первичная нейруляция - период 3-4 недели гестации; 2)Вентральная индукция - период 5-6 недели гестации; 3)Нейрональная...
-
Отношение растений к свету - Строение животных и растений
Как экологический фактор внешней среды свет прежде всего необходим для процесса фотосинтеза, в результате которого формируется 90-95 % органической массы...
-
Искусственный отбор - Сущность эволюционной теории Чарльза Дарвина
Исходным положением учения Дарвина является его утверждение о наличии изменчивости в природе. Изменчивостью Называют общее свойство организмов...
-
Большое значение в формировании эволюционных взглядов Дарвина сыграли его собственные открытия, сделанные им во время путешествия на корабле "Бигль"....
-
БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ - Сорные растения
Для успешной борьбы с сорняками необходимо знать их биологические особенности и способы распространения. За долгий период своего существования среди...
-
Введение, О лекарственных растениях и травах - Мир лекарственных растений
Мир растений - это величайшее чудо природы, наше целительное богатство и царство красоты. Каждое растение представляет собой своеобразную фабрику, в...
-
Роль микоризы в жизни растений Микориза - неотъемлемая часть большинства растений в природе. На инфицированных корнях грибы развивают внешний мицелий,...
-
Определено расположение субъединиц в геномах вирусов, прокариотов и эукариотов, послужившее основой для создания универсального генетического кода....
-
Омемла-- вечнозеленое кустарниковое растение, род полупаразитных кустарников. Паразитирует на очень многих древесных растениях: тополях, кленах, соснах,...
-
Семенная кожура - Покровные ткани растений
Особым типом покровной ткани является семенная кожура или семенная оболочка, которая образуется из покровов (интегументов) семязачатка и является...
-
Амая многочисленная группа облигатных подземных паразитов -- заразиховые. Род заразиха -- Оrobanche по сравнению с другими родами семейства отличается...
-
Город как комплекс природных и урбаноэкосистем и роль растений в формировании городской среды В современном мире стремительно идет процесс урбанизации. С...
-
Первые этапы развития анатомии в России в XVIII веке - История развития отечественной анатомии
Петр I всю жизнь испытывал огромный интерес к медицине, поэтому за границей он много времени уделял посещениям известных в то время врачей и...
-
Из всех перечисленных типов питания углеродом фотосинтез зеленых растений, при котором построение органических соединений идет за счет простых...
-
Аденозинтрифосфорная кислота (АТФ) состоит из органического основания аденина (I), углевода рибозы (II) и трех остатков фосфорной кислоты (III)....
-
Применение растения в народной методике лечения - Рожденный в Южной Америке
Цветок Амаранта Множественные народные рецепты рекомендуют применять растение для устранения грибковых заболеваний и шрамов, лечения экземы и герпеса, а...
-
История культуры. - Экологические группы растений по отношению к свету, воде и теплу
Гранат претендует на одно из первых мест в самых ранних анналах истории, поскольку по своему потребительскому значению он занимал у первобытных людей...
-
Применение - Экологические группы растений по отношению к свету, воде и теплу
Лечебные свойства алоэ, судя по сохранившимся письменным источникам, были известны уже более трех тысяч лет назад В настоящее время различные препараты...
-
Рассмотрим сначала питание растительноядных рыб. В Казахстане к ним, прежде всего, следует отнести белого амура, вывезенного с Дальнего Востока и...
-
Одним из аргументов против роли вирусов в возникновении большинства злокачественных опухолей у человека считается тот факт, что в подавляющем большинстве...
-
Какие свойства делают растение полезным - Рожденный в Южной Америке
Научные исследования подтверждают высокое содержание в растении белков, наличие витаминов (витамины А, С, D и Е и группа В) и минеральных веществ (до 4%,...
Этап. Филогенетическая (эволюционная) систематика - Систематика растений