Обоснование параметров и формы технологической комбинированной перегородки двухблочного пчелиного улья
Введение
Величайший пчеловод нашей эпохи П. И. Прокопович сказал: "Управление пчелами есть знание содержать каждое пчелиное семейство в надлежащем состоянии... словом, знать и уметь все производить в каждом улье с пчелами искусно, по науке, чтобы по возможности способствовать лучшим выгодам, доставляемым пчелиными семействами".
Если интерпретировать все сказанное современными терминами, то "знать и уметь все производить в каждом улье с пчелами искусно, по науке" и есть суть современных технологий интенсивного пчеловождения. А конечная цель - "способствовать лучшим выгодам, доставляемым пчелиными семействами" - есть суть рентабельного (рационального) пчеловодства [1].
Анализ источников
Все сложное проявление жизнедеятельности пчел направляется их нервной системой. Поведение пчел определяется врожденными, передающимися из поколения в поколение инстинктами и безусловными, а в некоторых случаях приобретенными в течение жизни условными рефлексами. Пчела имеет высокоразвитую нервную систему и органы чувств, благодаря которым осуществляется постоянная связь между особями пчелиной семьи и с внешней средой. Органами чувств пчела воспринимает изменения в окружающей обстановке. Сигнал об этом передается в центральную нервную систему, которая согласует работу мышц с изменениями внешних условий. Так происходит ответная реакция организма на раздражения [14].
Тело всех особей пчелиных семей снаружи покрыто кутикулой, которую называют хитином. Кутикула - продукт выделения подкожного слоя клеток эпидермиса, содержит около 50 % хитина (азотсодержащий полисахарид) и нерастворимые белковые вещества. В кутикуле различают два слоя: тонкий наружный - кутикулин и внутренний толстый - эндокутикулу. Наружные покровы предохраняют внутренние органы от высыхания и воздействия химических веществ. Тело покрыто волосками. Одни из них защищают от пыли, другие служат органами осязания [13].
Единство пчелиной семьи поддерживается комплексом взаимосвязей между ее членами. К ним относятся трофические и тактильные контакты (обмен кормом и феромонами), сигнальные звуки, движения, запах и др.
Феромоны пчел - биологически активные вещества, секретируемые железами насекомого в окружающую среду. Они имеют непосредственное отношение к поддержанию единства, регулированию физиологического состояния и поведению членов семьи. У пчел обнаружено до 30 различных феромонов, отличающихся между собой по специфичности физиологического воздействия на членов семьи. Из числа обнаруженных феромонов идентифицировано и синтезировано более десяти. Детально изучены феромоны матки [3, 12].
Еще в 1954 г. Батлер открыл в организме пчелиной матки "маточное вещество". Через пять лет удалось выделить его из головок пчелиных маток и определить строение. А еще через два года Батлер, Келлоу и Джонстон в выделениях мандибулярных желез головы пчеломатки нашли другое специфическое вещество, которое они назвали "запах матки" [2].
Наиболее высокой активностью и широким спектром действия обладает маточное вещество, так называемый феромон № 1 (транс-9-кето-2-деценовая кислота), продуцируемый верхнечелюстными железами пчеломатки. С его помощью пчеломатка привлекает трутней во время брачного полета в воздухе и рабочих пчел внутри пчелосемьи, а также в дозах 0,13-0,50 мкг [2] оказывает стерилизующее действие на рабочих пчел, предупреждая откладку ими неоплодотворенных яиц, и сдерживает выращивание в семье новых пчеломаток. Феромон № 1 в сочетании с исходящими от пчеломатки ароматическими соединениями, которые получили групповое название феромон № 2, оказывает регулирующее действие на рабочих пчел. В состав феромона № 2 входят метилфенилацетат (метиловый эфир фенилуксусной кислоты) и метилпропионат (метиловый эфир пропионовой кислоты). В секрете верхнечелюстных желез найден роестабилизирующий феромон (транс-9-окси-2-деценовая кислота). Он оказывает регулирующее воздействие в период роения пчел.
Рабочие пчелы, в окружение которых попадает пчеломатка при свободном передвижении по сотам, слизывают с ее тела феромоны и вместе с кормом передают остальным членам семьи. Через феромоны пчелы узнают о состоянии пчеломатки. При неудовлетворительном состоянии пчеломатки пчелы ее меняют. Гибель пчеломатки вызывает нарушение всей деятельности семьи независимо от количества в ней пчел: они прекращают строить соты, слабо работают на медосборе, плохо защищают свои гнезда. Пчелосемьи, оставшиеся без пчеломатки, часто покидают свои ульи.
Феромоны, имеющие отношение к регулированию физиологического состояния пчелосемьи, выделяют не только пчеломатка и другие взрослые особи, но и расплод. У рабочих пчел при скармливании им экстрактов из личинок подавлялось развитие яичников.
Помимо феромонов, связанных с размножением, в семье пчел существуют феромоны, регулирующие выполнение работ, необходимых для сохранения общественного образа жизни. Для охраны пчелосемьи от врагов существуют феромоны тревоги и отпугивания, выделяемые жалоносным аппаратом (изоамилацетат) и мандибулярными железами рабочих пчел (гептанон-2). Его содержание в одном жале составляет около 1 мкг. Запах такого изоамилацетата при 20 0С пчелы ощущают в течение 10 мин. [2, 3, 13].
Также установлено, что пчелы ориентируются и подают сигналы запахом - продуктом деятельности так называемых насоновых желез. Эти кожные железы пчелы (имеющие форму валика) называются также "пахучими". Секрет, который они выделяют, имеет специфический кислый запах.
Они имеются у пчеломатки и рабочих пчел и расположены между 5-м и 6-м тергитами брюшка. У рабочих пчел железа функционирует при доставке воды или в тех случаях, когда они посещают источники пищи, не обладающие запахом. Пчеломатка использует железу при вылетах из улья. Пахучие вещества железы служат ориентиром для пчел, сопровождающих пчеломатку во время роения [3].
А. Рут, К. Фриш, И. Халифман и другие ученые полагают, что каждой пчелиной семье свойственен свой, специфический, ульевой, "фамильный" запах. Наблюдения показали, что пчелиная стража, которая зорко охраняет леток, безусловно, пользуется обонянием для распознавания и пропуска своих пчел в пчелиный город. "Запах семьи" для нее лучший пароль. Этот запах служит пчелам компасом в полетах за нектаром, пыльцой и для безошибочного возвращения домой [4].
К. Фриш по этому поводу писал в своих работах: "У каждой рабочей пчелы имеется всегда наготове "маленький флакончик духов". Вблизи кончика брюшка со спинной стороны имеется складка в коже, которая завернута внутрь и поэтому невидима, но которая может произвольно выпячиваться в виде влажно-блестящего валика. При его выпячивании особыми железами выделяется в этот мешочек вещество, распространяющее сильный лимонный запах, характерный для известного медоносного растения мелиссы. Даже для человеческого обоняния этот запах явно ощутим. Этот запах и позволяет пчелам находить свой улей [4, 5].
Важное значение в жизни пчел занимают условные рефлексы, направленные на коллективное и эффективное использование появившегося в природе медосбора. Условные рефлексы играют большую роль в жизни пчелиной семьи: при ориентировочном облете пчелы запоминают место расположения своего улья, при полете на медосборе - путь к источнику нектара и пыльцы и обратно к пасеке; запоминают окраску цветков, их форму и запах.
Условные рефлексы приобретаются сравнительно быстро, но также быстро утрачиваются. Если улей перевезти на 5-7 километров, то пчелы, проведя ориентировочный облет, утратят рефлекс ориентации на старое место. С прекращением выделения нектара пчелы утрачивают рефлекс на цвет, форму и запах цветков, с которых они брали нектар [14].
Используя систему акустической сигнализации, в качестве линий акустического канала связи пчелы используют воздух или твердые тела. В принципе любой звуковой сигнал, независимо от способа генерации, может передаваться по каждой из этих линий. Однако с точки зрения экономии энергетических затрат акустические сигналы целесообразно передавать, минуя переходы через среды с различной акустической проводимостью, что, по-видимому, повлияло в филогенезе на организацию системы акустической сигнализации пчел. Действительно, они обычно используют воздушную среду в тех случаях, когда звук генерируется без участия твердого опорного субстрата. Твердый субстрат как линию связи пчелы используют для передачи сигналов, генерируемых посредством воздействия на субстрат. Это достигается тем, что звукоиздающий орган контактирует с субстратом.
Воспринимают пчелы сигналы, передаваемые по воздуху и через субстрат, двумя различными типами рецепторов. Роль рецепторов, воспринимающих колебания субстрата, выполняют субгенуальные органы, а распространяющихся через воздушную среду, - специализированные фонорецепторы, расположенные на голове [15].
Механические раздражения пчелы воспринимают с помощью хордотональных органов, представляющих собой группу особых сенсилл, натянутых между двумя участками кутикулы. Натянутые, как струны, они регистрируют малейшие деформации и воспринимают степень натяжения стенки тела. Эти сенсиллы, наряду с функциями проприорецепторов, нередко приобретают способность к регистрации звуков. Хордотональная сенсилла состоит из трех клеток: колпачковой, обкладочной и чувствительного нейрона. Колпачковая и обкладочная клетки равноценны двум эпидермальным клеткам осязательного волоска. Отличительная черта хордотональных сенсилл - наличие канала, в котором заключен наружный отросток чувствительной клетки.
Еще одной специализированной формой хордотональных органов является Джонстонов орган. Он расположен на втором членике антенны медоносной пчелы.
Сколопофоры Джонстонова органа образуют в педицеллярном членике цилиндр вокруг осевых нервов антенны и реагируют на смещение третьего членика относительно второго. Стимуляция Джонстонова органа происходит как при активном ощупывании антеннами различных предметов, так и во время движения насекомого в воздушной среде. Джонстонов орган, кроме того, воспринимает звуковые колебания. Пчелы способны воспринимать не только звуковые колебания, но и любые колебания среды. Если ухо человека воспринимает изменения давления воздуха, вызываемое источником звукового колебания, то пчелы чаще воспринимают движение воздуха.
В естественных условиях органы слуха могут реагировать на всевозможные звуки как биотического, так и абиотического характера.
Однако в первую очередь медоносные пчелы приспособлены к восприятию лишь тех звуковых сигналов, которые имеют для них коммуникативное значение [16].
Итак, мы выяснили, что каждая пчелиная семья имеет свои индивидуальные особенности: агрессивность, способность к сбору меда, прополисованию гнезд, зимостойкость, ройливость, - но основными и отличающими являются специфический запах и звук, которые сохраняются лишь до тех пор, пока в ней живет одна и та же матка.
Методы исследования
Методы исследования - дедуктивно-экспериментальный, на основании которого проводились биологические, физико-математические исследования движения естественного воздушного потока и свободное проникновение индивидуального запаха и звука одной пчелиной семьи в объем ульевого блока, занимаемого второй семьей, и наоборот. Информационной базой послужили труды отечественных и зарубежных исследователей, научные разработки, данные экспериментов, полученные в результате наблюдений, опытов и расчетов авторов.
1. Основная часть
Присматриваясь к общению пчел, разных семей с присущим только для них индивидуальным запахом и звуком, можно узнать многое об их жизни; уберечь пчеловода от их агрессивного настроения, избежать гибели подсаживаемой матки, привлечь рой, переставить улей на новое место, объединить две пчелосемьи в двухблочном пчелином улье с целью создания одной более сильной с двумя пчеломатками.
Рассматривая биологические особенности пчелиной семьи, видим, что интенсивность сбора углеводного корма в ней находится в прямой зависимости от общего количества пчел [6]. Говоря о сильной пчелиной семье, предполагаем 60-80 тысяч пчел, т. е. 6-8 кг. При этом можно утверждать, что такая сильная семья за время главного медосбора при хорошей кормовой базе может собрать до 100-150 кг меда. Обычно такие семьи получаются в результате объединения нескольких роев: естественным путем образуется свальный рой, искусственным их соединением - рой ссыпчак. Однако о биологической целостности и гармоническом составе таких пчелосемей говорить нельзя. В результате в такой сильной пчелиной семье с одной пчеломаткой количество выделяемого ею маточного вещества явно недостаточно для всех рабочих особей. По этой причине такие семьи проявляют тенденцию к немедленному роению, закладке свищевых маточников, а иногда даже к элементарному изгнанию части рабочих пчел [1]. Следовательно, получить пчелиные семьи с одной пчеломаткой к главному медосбору естественным путем в нашей природно-климатической зоне с широкоиспользуемым качеством пчеломаток физически невозможно. Но выход есть - мирное объединение рабочих особей соседних пчелосемей, каждая из которых имеет свою плодную пчеломатку и готова максимально эффективно использовать медосбор любой силы.
Для осуществления этой цели нами предложена технологическая комбинированная перегородка, основной ролью которой является полное исключение времени на конфликты и гибель между особями соседних пчелиных семей двухблочного пчелиного улья в период заполнения общего магазинного корпуса нектаром. Первый технический результат указанной перегородки - свободное проникновение индивидуального запаха и звука одной пчелиной семьи в объем основного корпуса, занимаемого второй семьей, и наоборот. Обмен запахом пчелиных семей происходит через объединяющее окно, представляющее собой зарешеченный проем в комбинированной технологической перегородке. Параметры объединяющего окна определены экспериментально-опытным путем. Нами исследовано три наиболее вероятных типа-размера объединяющего окна технологической комбинированной перегородки (рис.1 а, б, в).
![технологическая комбинированная перегородка двухблочного пчелиного улья [10]](/images/image001-1201.jpg)


Рис. 1 Технологическая комбинированная перегородка двухблочного пчелиного улья [10]: а) объединяющее окно занимает всю площадь рамки, б) объединяющее окно занимает 1/2 площади рамки, в) объединяющее окно занимает 1/4 площади рамки:1 - нижний брусок, 2 - объединяющее окно, 3 - двухсторонний экран, 4 - две боковые планки, 5 - верхний брусок
Оптимальный размер объединяющего окна (100Ч290Ч25) мм, расположенного симметрично вертикальной оси перегородки, указали сами пчелы, так как остальную зарешеченную поверхность в 70 % случаев просто прополюсуют. Вероятно, при увеличении размеров объединяющего окна увеличивается скорость свободного воздушного потока - возможен сквозняк, что недопустимо. Уменьшение размеров нецелесообразно потому, что обоюдно проникающий индивидуальный запах семей не полностью охватит соответствующий объем и особи, находящиеся на дальних рамках от перегородки, не ощутят проникающего запаха, и для них он останется неизвестным более продолжительное время. Выбор размера ячейки сетки основан исходя из биологических свойств пород пчел, используемых на пасеках нашей страны, и составляет не более 9 мм2 [17].
Нами предлагается установка технологической комбинированной перегородки во втором корпусе двухблочного пчелиного улья, корпус при этом изготавливается на стандартную, наиболее распространенную в нашей стране рамку Дадана с разделителями, отсюда вытекает выбор формы (рис. 2) [7]. Однако с учетом основной роли технологической комбинированной перегородки габаритные размеры ее должны быть иными (рис. 3).
Рис. 2 Стандартная рамка Дадана с разделителями
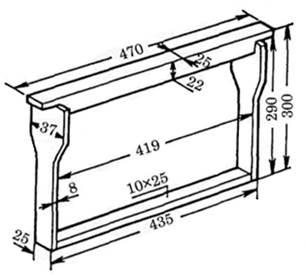

Рис. 3 Габаритные размеры и форма экспериментальной технологической комбинированной перегородки
Известно, что размеры между боковыми планками рамки и стенками улья составляют 7 мм, а расстояние между нижним бруском рамки и верхним срезом гнездового корпуса равно 20 мм. Определим габаритные размеры рассматриваемой перегородки.
Для удобства работы с технологической комбинированной перегородкой, приняв во внимание допуски на материал, применяемый для ее изготовления, определим размеры:
- - по ширине: 435 мм + 5 мм + 5 мм = 445 мм; - по высоте: 300 мм + 19 мм = 319 мм.
Таким образом, общая площадь технологической комбинированной перегородки равна:
445 Ч 319 = 141955 мм2.
Определение толщины технологической комбинированной перегородки не является столь простой задачей. Для этого вновь вернемся к биологии пчелы. Ротовой аппарат пчелы относится к грызуще-сосуще-лижущему типу. Он представлен двумя независимыми органами, образующими грызущую и сосущую системы. К грызущей системе относятся мандибулы и верхняя губа, к сосущей - максилла (нижняя челюсть) и нижняя губа. Нижние челюсти совместно с нижней губой образовали вытянутый хоботок [11]. У пчелиной матки мандибулы более широкие и на конце имеют острый зубец, которым она надрезает восковую крышечку перед выходом из маточника. Мандибулы трутня недоразвиты [12]. Мандибулами рабочая пчела выполняет несколько действий: разрывает пыльники на цветках при сборе пыльцы, разминает воск при отстройке сотов, разгрызает дерево при расширении очень узкого летка, схватывает соринки и выносит их из улья, хватает других пчел, врагов [13]. Строение хоботка у всех особей пчелиной семьи одинаковое. Разница заключается только лишь в его длине. Длина хоботков у маток и трутней различных пород пчел почти одинакова. У маток среднерусских пчел она равна 4,14 мм, у трутней - 4,04 мм, итальянских - 4,06 и 4,08 мм, краинских - 4,24 и 4,48 мм и кавказских - 4,35 и 4,23 мм соответственно. Наиболее длинный хоботок у рабочих пчел, но его длина колеблется в больших пределах, чем у маток и трутней, в зависимости от принадлежности к той или иной породе - от 5,5 до 7,2 мм [12]. На основании этого нами проведены исследования, результаты которых отображены графически по годам (рис. 4). Длинна хоботка, экстерьер и другие биологические признаки наиболее распространенных пород пчел представлены в таблице [9].


Рис. 4 Зависимость гибели пчелиных маток и рабочих особей соседних пчелосемей от толщины технологической комбинированной перегородки двухблочного пчелиного улья: а) активный период 2010 г., б) активный период 2011 г., в) активный период 2012 г.
Экстерьер и биологические признаки пчел наиболее распространенных пород
За длину хоботка принимают расстояние от вершины подбородка до наружной границы ложечки нижней губы [11]. Принимая во внимание тот факт, что в корпусе, где устанавливается указанная технологическая перегородка, используются рамки Дадана с толщиной верхнего бруска в 25 мм, а это полностью удовлетворяет требованию параметра по толщине используемой технологической перегородки, то принимаем ее размер, равный 25 мм (рис. 5).
|
Порода |
Основные признаки |
Косвенные признаки | ||||||||||
|
Окраска тела пчелы |
Длина хоботка, мм |
Кубит. индекс % |
Ширина третьего тергита, мм |
Печатка меда |
Зимостойк. |
Поведение пчел |
Масса, мг |
Плод-сть матки яиц/сут. | ||||
|
При осмотре сота |
При открывании гнезда |
Однодн. пчелы |
Неплод. матки |
Плодн. матки | ||||||||
|
Среднерусская |
Темно-серая |
6,0-6,4 |
60-65 |
4,8-5,2 |
Белая |
Хорошая |
Покидают сот |
Агрессивное |
110 |
190 |
210 |
1500-2000 |
|
Серая горная кавказская |
Серая |
6,7-7,2 |
50-55 |
4,4-5,0 |
Темная |
Слабая |
Сидят на соте |
Миролюбивое |
90 |
180 |
200 |
1100-1500 |
|
Желтая кавказская |
Желто-серая |
6,5-6,9 |
50-55 |
4,4-5,0 |
То же |
То же |
То же |
То же |
90 |
180 |
200 |
1100-1700 |
|
Карпатская |
Серая |
6,3-7,0 |
45-50 |
4,4-5,1 |
Белая |
Удовлетв. |
То же |
То же |
110 |
185 |
205 |
1100-1800 |
|
Украинская степная |
Серая |
6,3-6,7 |
55-60 |
4,6-5,1 |
Белая |
То же |
То же |
Беспокойное |
105 |
180 |
200 |
1100-1800 |
|
Краинская |
Серая |
6,4-6,8 |
45-50 |
4,7-5.1 |
То же |
То же |
Передвиг по соту |
То же |
110 |
185 |
205 |
1400-2000 |
|
Итальянская |
Желтая |
6,4-6,7 |
40-45 |
4,7-5,2 |
Смешан. |
Слабая |
То же |
То же |
115 |
190 |
210 |
1500-2500 |
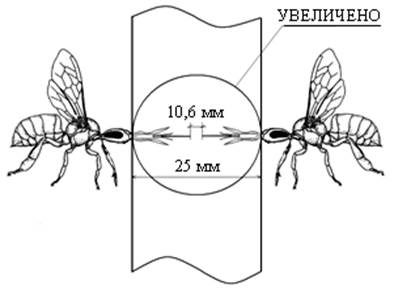
Рис. 5 Фрагмент технологической комбинированной перегородки с гарантированным зазором, предохраняющим от контакта хоботками пчелиных особей соседних семей
Пчелиные матки и рабочие особи, встречаясь у объединяющего окна 2 (рис. 1 в), лишены возможности физического контакта, так как сетка окна с ячейкой 9 мм2 препятствует их встрече, а толщина перегородки 9 (рис. 5) в 25 мм исключает контакт хоботками, в результате матки и рабочие особи соседних пчелосемей не могут смертельно травмировать друг друга и семьи мирно существуют [8, 9].
К числу не менее важных средств, имеющих отношение к объединению пчелиных семей в биологическую единицу, относится и звук, издаваемый пчелами и пчеломатками. Остановимся только на анализе и характеристике роли совокупного общего звука, издаваемого пчелами, т. е. звуковом фоне пчелосемьи. [15] Общий звук семьи, который слагается из гула тысяч пчел, можно всегда слышать, приложив ухо к улью или с помощью обыкновенного стетоскопа. Звуки, издаваемые пчелами, состоят из звуковых волн разной частоты. Например, звуки, образуемые пчелами при входе и выходе из улья, имеют частоту около 180 герц. Незадолго до начала роения в семье появляются пчелы-кормилицы с избытком маточного молочка, они издают звуки уже 25530 Герц. При подготовке пчелосемьи к роению появляется совершенно другой звук, высотой свыше 3000 Герц [13].
Спектр фоновых составляющих, отличающихся по интенсивности на несколько десятков децибел, лежит в широком диапазоне частот. Низкочастотные составляющие звукового фона граничат с инфразвуковой областью, высокочастотные находятся в диапазоне 8000-12000 Герц. Максимальная энергия фона сосредоточена в трех диапазонах, порождаемых соответствующими процессами жизнедеятельности каждой пчелиной семьей. Первый диапазон находится в пределах 70-180 Герц, второй-- 200-400 Герц, третий - 400-550 Герц. [15]. Первый диапазон интенсивных составляющих фона порождается звуками, возникающими при вентилировании улья, на что указывает прежде всего совпадение этой области спектра с областью интенсивных составляющих в звуках пчел-вентилировщиц. Интенсивность звуков в этом диапазоне частот находится в прямой зависимости от уровня активности пчел-вентилировщиц и потому в годовом цикле минимальна в период зимовки, когда пчелы более пассивны. В этот период интенсивность составляющих первого диапазона на 9-14 Дб. ниже, чем во втором. В летние месяцы жизни пчелосемьи интенсивность звуков, издаваемых пчелами-вентилировщицами, достигает, а иногда и превосходит интенсивность составляющих второго диапазона в спектре фона. Интенсивность первого диапазона зависит в этот период от внешней температуры и количества доставляемого в улей нектара. С повышением температуры или увеличением количества нектара возрастает активность пчел-вентилировщиц и соответственно увеличивается интенсивность издаваемых ими звуков [15].
Второй диапазон интенсивных составляющих спектра фона улья порождается звуками, сопровождающими работу пчел по обогреванию гнезда, на что указывает прямая связь между интенсивностью этого диапазона и количеством тепла, выделяемого пчелосемьей. К тому же установлено, что пчелы могут издавать звук в этом частотном диапазоне исключительно вибрацией торакса под действием мышц непрямого действия, а вибрация торакса этими мышцами сопровождается значительным выделением тепла. Интенсивность составляющих второго диапазона спектра прямо связана также с уровнем двигательной активности пчел, наблюдаемой и в тех случаях, когда семья не нуждается в обогреве. Генерировать звуки с максимальной интенсивностью на частотах до 600 Герц пчелы способны только потому, что торакс вибрирует под действием продольных мышц, которые способны сокращаться в высоком ритме (рис. 6). Из рисунка 6 видно совпадение частоты движения крыльев в летательной плоскости с несущей частотой звуковых сигналов подтверждает преимущество продольных мышц в издавании пульсирующих звуков - это подтверждают осциллограммы вибраций по Е. Еськову [15].
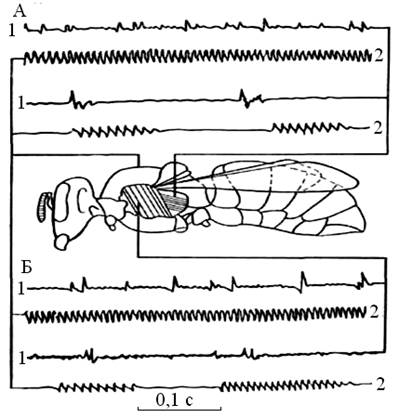
Рис. 6 Осциллограммы: 1 - потенциал действия, 2 - вибрации торакса, А - продольные мышцы, Б - поперечные мышцы
Соотношение интенсивностей составляющих второго диапазона спектра фона зависит от физиологического состояния и уровня активности пчел. В летний период жизни пчел положение пика интенсивности в этом диапазоне во многом определяется уровнем активности пчел, занятых доставкой в гнездо нектара и пыльцы. Эта зависимость выражается в том, что при понижении активности пчел-сборщиц пик интенсивности смещается на низкочастотную область рассматриваемого диапазона, а при повышении - на высокочастотную.
Колебания температуры наружного воздуха, влияющие на изменение общего уровня интенсивности звуков пчелиной семьи, соответственно изменяют уровень составляющих в диапазоне 200-400 Герц. Однако частотное положение пика интенсивности не зависит от температуры среды, если активность пчел-сборщиц находится на одном уровне. Смещение пика интенсивности на низкочастотную область второго диапазона происходит лишь при понижении температуры до уровня, тормозящего летную активность пчел. На соотношение интенсивностей составляющих второго диапазона существенно влияют изменения физиологического состояния пчел улья. В суточной и сезонной динамике спектральной структуры звукового фона также прослеживается связь с уровнем активности пчел.
В суточном цикле эта связь выражается в изменениях частотного положения пика интенсивности во втором диапазоне интенсивных фоновых составляющих. В дневные часы пик интенсивности находится в высокочастотной области этого диапазона, а ночью смещается на низкие частоты. Периоды повышения активности пчел в ночное время, стимулируемые охлаждением гнезда, отражаются на повышении частоты максимально интенсивной спектральной составляющей второго диапазона [15].
Связь между структурой спектра в диапазоне 200 - 400 Герц и уровнем активности пчел в сезонном цикле выражается в основном с изменением частотного положения пика интенсивности. В период активной жизни пчел этот пик находится в области высоких частот второго диапазона, а в зимние месяцы смещается на его низкочастотную область. Узкие полосы спектрального фона свидетельствуют о том, что он имеет пульсирующую структуру. По частоте пульсаций в спектре фона четко различаются три частотные области, соответствующие по положению трем диапазонам, в которых сосредоточена основная энергия звуков пчелиной семьи. Частота пульсаций фона мало отличается в семьях пчел разных рас. Однако изменение активности пчел в летний период влияет на структуру звукового фона улья. Сильное возбуждение пчел, например, под воздействием электрического поля понижает частоту пульсаций фоновых составляющих, в период спада активности, следующим за отключением поля, частота пульсаций фона превосходит исходный уровень. Это указывает на наличие обратной связи между частотой пульсаций, фоновых составляющих и уровнем активности пчел.
Рассматривая звуковые процессы с точки зрения использования их пчелами как средства координации их деятельности, можно выделить следующее. Дискретная структура частотно-амплитудного спектра звукового фона пчелиной семьи обусловлена, вероятно, процессом взаимной подстройки звучания пчел. Это, наряду с синхронизацией ритма пульсаций в различных диапазонах, является одним из проявлений общей закономерности автосинхронизации процессов, протекающих в биологических системах. Такая синхронизация звуков, издаваемых пчелами улья, указывает на то, что в семье нет какого-то регулирующего центра. Тем не менее автосинхронизация звуковых процессов в пчелиной семье способствует объединению ее членов в некоторую взаимосвязанную и взаимозависимую биосистему [15].
Все это подтверждает факт взаимодействия семей и влияния "настроения" одной пчелиной семьи на другую, находящуюся за звукопроницаемой технологической комбинированной перегородкой. Следовательно, звуки также играют определенную роль при объединении рабочих особей соседних пчелосемей двухблочного пчелиного улья в единую сверхсильную пчелиную семью.
Для наглядности рассуждений на рис. 7 указано место установки технологической комбинированной перегородки 9 в двухблочном пчелином улье, который состоит из гнездового корпуса 6, основного корпуса 7, магазинного корпуса 8 [10].
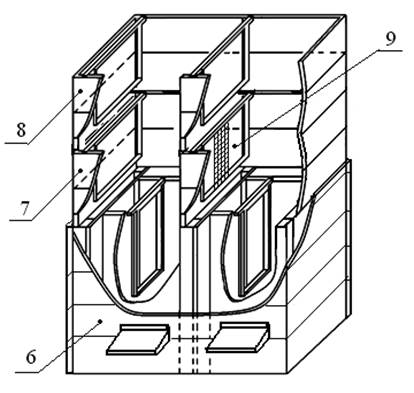
Рис. 7 Место установки технологической комбинированной перегородки в двухблочном пчелином улье
Заключение
Проведенные нами исследования показали, что, используя биологические особенности пчелиных семей, возможна установка технологической комбинированной перегородки и получение ожидаемого технического результата. Так, при установке в основной корпус двухблочного пчелиного улья комбинированной технологической перегородки через объединяющее окно происходит свободное проникновение индивидуального запаха и звукового фона одной пчелиной семьи в объем, занимаемый второй семьей, и наоборот. При этом физический контакт пчелиных маток и рабочих особей соседних пчелосемей до времени приобретения нового запаха и звука полностью исключен. В итоге при заполнении общего магазинного корпуса нектаром полностью исключается возможность конфликтных ситуации между пчеломатками и рабочими особями обеих семей, так как запах и звуковой фон соседней семьи уже знаком. Это повышает производительность объединенной пчелиной семьи на 8-12 % и более, в зависимости от климатических условий и кормовой базы.
Литература
- 1. Корж, В. Н. Пчеловодство. Практический курс / В. Н. Корж. - Ростов н/Д: Феникс, 2008. - 543.с. 2. Машенков, О. Н. Чем пахнут пчелы / О. Н. Машенков // Пчеловодство. - 2002. - №1. - С 18-19. 3. Харчук, Ю. Справочник по домашнему пчеловодству / Ю. Харчук. - [Электронный ресурс]. - Режим доступа: http://lib. rus. ec/ (fb2) - Дата доступа: 11..05.2012. 4. "Запах семьи" у пчел. - [Электронный ресурс]. - Режим доступа: http://www. happy-giraffe. ru/community/34/forum/post/17096/ - Дата доступа: 10.05.2012. 5. Фриш, К. Из жизни пчел / К. Фриш; перевод с немецкого Т. И. Губиной; под ред. И. А. Халифмана. - М.:"Мир", 1980. - 212 с. 6. Шершнев, А. Н. Оптимальная сила пчелиной семьи и ее расчет / А. Н. Шершнев // Вестник БГСХА. - 2010. - №4. - С. 102-105. 7. Основы пчеловодства: учеб. пособие / В. М. Каплич [и др.]. - Минск: БГТУ, 2009. - 408 с. 8. Дударевич, М. А. Объединение - дело тонкое / М. А. Дударевич // Хозяин. - 1999. - № 4. - С. 22-24. 9. Медвецкий, Н. С. Пчеловодство / Н. С. Медвецкий. - Минск: Ураджай, 2000. - С. 217. . 10. Шершнев, А. Н. Определение объема двухблочного пчелиного улья / А. Н. Шершнев // Вестник БГСХА. - 2012. - №2. - С. 115-119. 11. Риб, Р. Д. Методика оценки экстерьерных признаков / Р. Д. Риб // Пчеловодство. - 2009. - № 7. - С. 64-66. 12. Козин, Р. Б. Биология медоносной пчелы: учеб. пособие / В. И. Лебедев, Н. В. Иренкова - СПб.: Издательство "Лань",2007. - 320 с. 13. Таранов, Г. Ф. Анатомия и физиология медоносных пчел / Г. Ф. Таранов. - М.: Колос, 1968. - 344 с. 14. Котова, Г. Н. Биология пчелиной семьи / Г. Н. Котова. - М.: Россельхозиздат, 1974. - 34 с. 15. Еськов, Е. К Поведение медоносных пчел / Е. К. Еськов. - М: Колос, 1981. - 183 с. 16. Кривцов, Н. И. Пчеловодство / Н. И. Кривцов, В. И. Лебедев, Г. М. Туников. - М.: Колос, 2007. - 512 с. 17. Шеметков, М. Ф. Справочник пчеловода / М. Ф. Шеметков. - Минск: Урожай, 1969. - 467 с.
Похожие статьи
-
Биология пчелиной семьи - Пчеловодство
Пчела медоносная вместе с осами, муравьями, шмелями и некоторыми другими формами относится к общественным насекомым. Структура сообщества медоносной...
-
Хлебная масса с поля поступает на рабочие органы, производительность которых должна быть согласована между собой. Однако изменения условий уборки...
-
Комплексная оценка и отбор пчелиных семей для разведения - Разведение с основами частной зоотехнии
Существующие в настоящее время рекомендации по племенной работе в пчеловодстве разработаны в большинстве своем по аналогии с другими отраслями...
-
Оценка зимостойкости пчелиных семей - Разведение с основами частной зоотехнии
Зимостойкость - важнейший качественный показатель пчелиных семей, с которого нужно начинать и которым следует заканчивать работу по чистопородному...
-
В данном курсовом проекте для выполнения основных работ на лесосеке применяются укрупненные комплексные бригады (УКБ) и функциональные бригады (звенья)....
-
Тип технологического процесса, состав, последовательность и место выполнения операций лесосечных работ определяются видом продукции, погружаемой на...
-
Ветеринарно-гигиеническое требования к оценке территории фермы Для выбора земельного участка под строительство животноводческих предприятий, зданий и...
-
Обоснование вопроса Бурное развитие сельского хозяйства привели к увеличению продуктивности зернового хозяйства и скоплению на токах крупных колхозов и...
-
Организация пасеки - Пчеловодство
Пасеки. Пасекой принято называть небольшое пчеловодческое хозяйство, располагающее земельным участком с размещенными на нем временными или постоянными...
-
Организационно-производственная структура хозяйства - это сочетание внутрихозяйственных производственных подразделений и аппарата управления, которое...
-
Для обеспечения работы комбайна необходимо выполнение условия: N Дв ? N Т , (6.1) Где N Дв - мощность двигателя, КВт ; N Т - мощность, необходимая...
-
Коровник скот крупный рогатый Коровник на 100 голов двухрядный, разделен на стойла, в которых размещают по 25 голов (4 стойла). Здание одноэтажное....
-
Обоснование параметров регулирующей осушительной сети - Мелиорация участка переувлажненных земель
Регулирующая сеть служит для сбора и удаления с орошаемой территории избыточных поверхностных и грунтовых вод. С помощью регулирующей части системы...
-
Для обоснования конструктивно-режимных параметров манипулятора, нами были выполнены теоретические исследования его рабочего процесса. Так как точка...
-
Структура урожая показывает Из каких элементов складывается его величина, при какой доле их участие формируется запланированный урожай. Основные...
-
Продуктивность пчелиной семьи представляет собой суммарный результат производства семьями разных видов пчеловодной продукции. Но в практической...
-
В соответствии с зоотехническими требованиями каждый вид корма приводят в состояние, обеспечивающее наилучший эффект при его скармливании...
-
Выбор технологической схемы осуществляется с тенденцией развития отрасли. Выбранная схема предопределяет направление переработки отдельных видов сырья....
-
В динамическом отношении машинно-тракторный агрегат представляет собой систему твердых тел, связанных между собой жесткими и упругими связями. Агрегат...
-
Оценка развития пчелиных семей - Разведение с основами частной зоотехнии
Показатели развития и продуктивности пчелиных семей в большей степени, чем другие качественные характеристики, подвержены влиянию различных факторов. Это...
-
Культивация - агротехническое мероприятие, обеспечивающее крошение, рыхление и частичное перемешивание почвы, а также полное уничтожение сорняков и...
-
Программирование урожая с.-х. культур - разработка комплекса технологических приемов, обеспечивающего оптимизацию регулируемых факторов среды для...
-
Обоснование необходимости разработки При хранении сортового картофеля необходимо более строго соблюдать технологический режим, от этого зависит качество...
-
Организация производства в животноводстве основывается на объективном действии экономических законов социализма и некоторых других факторов, важнейшим из...
-
12) Для установления размера шаговых (технологических) групп свиней вначале рассчитываем соответствующую группу свиноматок и ремонтных свинок, отобранных...
-
В процессе естественного расширения пчелами ареала за счет расселения в зоны с умеренным и холодным климатом отбор благоприятствовал накоплению...
-
Описание бизнеса - Развитие пчеловодства
Для производства молока будет организовано предприятие, которое будет иметь называние "Литр молока". Организационно-правовая форма - ИП. пчеловодство...
-
Оценка поведения пчел - Разведение с основами частной зоотехнии
Под поведением пчел нужно понимать совокупность особенностей состояния и реакций отдельных особей и семьи в Целом, направленных на реализацию их...
-
Помещение для содержания овцематок на 850 голов (в тепляке минимум 15% - 128 голов) живой массой - 60 кг. Норма площади для овцематок - 1-1.2 м на голову...
-
1).Я выбираю челночный петлевой способ движения т. к. он Позволяет производить холостые движения агрегата с наименьшими потерями, например: повороты...
-
Выбор и обоснование технологии производства - Технологический процесс очистки зерна
В настоящее время продукты переработки растениеводческой продукции являются основой пищевого рациона человека. Зерно и продукты его переработки содержат...
-
Таблица 9 Наименование работ и их последовательность Оптимальные сроки выполнения работ Препараты, норма расхода Состав агрегата 1. Лущение стерни после...
-
Технологическая схема выращивания сеянцев и саженцев в питомнике (Форма 6) - Лесные культуры
Наименование работ Время проведения работ Состав агрегата Трактор Орудие Паровое поле (ранний пар) 1. Известкование почвы перед основной вспашкой весной,...
-
Назначение и сущность линии Интенсификация производства продуктов животноводства на современных сельскохозяйственных предприятиях сопровождается выходом...
-
Основная причина порчи рыбы - ферментативные процессы, ведущие к распаду тканей в результате глубокого автолиза, активной жизнедеятельности различной...
-
Животноводческий ферма навоз органический Грамотно организованная утилизация отходов очень важна для успешного ведения конкурентоспособного хозяйства, к...
-
На современном этапе развития методов генетики широкое применение нашли методы ДНК-маркирования, которые основаны на анализе полиморфизма генома....
-
Размер левад. Группа лошадей Левады Норма площади на 1 м Жеребцы Индивидуальные 600 Кобылы Групповые 20 Молодняк Групповые 20 В левадах, примыкающих к...
-
Подготовка к работе плуга: А)Агротехнические требования: Все виды вспашки проводят ( кроме перепашки зяби, пара и заделки органических удобрений) должны...
-
Обоснование проекта включает расчеты, определение и доказательство экономической эффективности изменений, вносимых в землепользование; установление...
Обоснование параметров и формы технологической комбинированной перегородки двухблочного пчелиного улья