Молекулярно-генетическая идентификация аллелей S2 и S10 гена самонесовместимости у кребов и элитных селекционных форм яблони
На современном этапе развития методов генетики широкое применение нашли методы ДНК-маркирования, которые основаны на анализе полиморфизма генома. ДНК-маркеры широко используются при изучении генофонда культурных растений: для оценки генетических взаимосвязей изучаемых образцов в коллекциях генетических ресурсов; для ДНК-паспортизации сортов; картирования генов и их отслеживания в селекционном процессе (маркер-опосредованная селекция, позволяющая проводить идентификацию и отбор генотипов, несущих целевые гены, без их оценки по фенотипу) [1]. Необходимо отметить, что ДНК-маркерный отбор может быть наиболее востребованным при оценке признаков, для которых фенотипическая оценка является сложной процедурой или требует временных затрат и зависит от условий окружающей среды.
Для яблони - основной плодовой культуры, высокой актуальностью обладает вопрос, связанный с подбором опылителей и, соответственно, связанным с этим определением совместимости сортов при опылении. От этого напрямую зависит продуктивность садового агроценоза. В системе самонесовместимости яблони основная функция в регуляции принадлежит гену самонесовместимости (S-ген). Продукты S-гена в пестике - цитотоксические протеины с рибонуклеазной активностью (S-РНКазы). В процессе самонесовместимого взаимодействия S-РНКазы контролирует прорастание пыльцевой трубки, проникая в цитоплазму и деградируя пыльцевую РНК [2]. Пыльцевые трубки, прорастающие из пыльцы с определенной аллелью S-гена, ингибируются в пестиках растений, несущих тот же аллель [3]. Следовательно, оплодотворение блокируется, когда S-аллель пыльцы совпадает с одной или двумя S-аллелями пестика. Два сорта, несущие идентичный набор аллелей S-гена являются несовместимыми. При взаимоопылении таких сортов оплодотворения и последующего формирования плодов не происходит. Сорта, частично совместимые, несут по одной одинаковой S-аллели. Они могут являться опылителями друг для друга, однако эффективность опыления при этом снизится на уровень около 50% [2]. Необходимо сказать, что данный уровень снижения будет обусловлен исключительно генотипом сорта, при воздействии дополнительных факторов (расхождения в сроках цветения, низкая жизнеспособность пыльцы сорта-опылителя, неблагоприятные погодные условия) уровень закладки плодов снизится еще значительнее. Очевидно, что для максимальной реализации потенциала продуктивности сортов яблони необходимо выполнять формирование сортовых схем садового насаждения с учетом совместимости при опылении - использовать наиболее эффективных опылителей или комбинации сортов с наибольшим уровнем совместимости. В этой связи, важным этапом является определение аллельного набора S-гена у сортов.
Одним из перспективных путей получения максимального уровня опыления в садовом агроценозе является использование так называемых кребов яблони - сортов и форм, представляющих, как правило, образцы видов рода Malus или их межвидовые гибриды с яблоней домашней (Malus domestica Borch). Ряд кребов уже зарекомендовали себя в качестве перспективных опылителей для создания моносортных насаждений промышленных сортов, однако, учитывая разнообразие данных сортов и форм и постоянное пополнения промышленного сортимента яблони новыми сортами, следует отметить важность знаний об аллельном составе S-гена у кребов. Это позволит рекомендовать наиболее перспективные среди них для сортов яблони.
Очевидно, что применение ДНК-маркеров для экспресс идентификации аллельных комбинаций данного гена обладает значительным преимуществом в сравнении с фенотипической оценкой совместимости, так как может быть выполнено в сжатые сроки. На настоящее время молекулярно-генетический контроль данного процесса глубоко изучен, известно порядка 25 аллелей S-гена. Однако в культурном мировом генофонде яблони наиболее распространены 7-8 основных аллелей, из которых аллели S2, S3, S5, S7, S10 являются наиболее распространенными [4, 5].
Использование ДНК-маркеров определило возможность идентификации аллелей гена самонесовместимости среди образцов коллекций генетических ресурсов яблони в различных странах, где возделывается данная культура [6]. В отечественных генетических исследованиях яблони также получены результаты об аллельных комбинация искомого гена у сортов российской селекции, а также перспективных селекционных форм (элитных форм) [7 - 10].
Важность этих исследований очевидна - появляется возможность прогнозировать степень совместимости сортов при опылении, что позволяет эффективно комбинировать сорта в садовом насаждении, размещая максимально совместимые сорта в саду в непосредственной близости. Это позволит получать наибольший уровень завязи плодов и, соответственно, урожайности, а также создавать моносортные насаждения, экономя затраты на проведение защитных мероприятий и уборку плодов. Наряду с этим, данные об аллельном составе гена самонесовместимости могут являться частью ДНК-паспортов сортов яблони в комплексе с микросателлитным ДНК-фингерпринтом.
В связи с вышесказанным, в задачи наших исследований входило выполнение ДНК - маркерной идентификации аллельных комбинаций гена самонесовместимости у сортов и форм кребов. На данном этапе работы целевыми являлись аллели S2 и S10, относящиеся к числу наиболее распространенных аллелей в мировом сортименте яблони.
Материал и методы исследований
Объектами исследования послужили кребы и элитные селекционные формы яблони. ДНК экстрагировали методом ЦТАБ [11]. Для идентификации аллелей гена самонесовместимости использовали метод полимеразной цепной реакции ПЦР, согласно общепринятым рекомендациям [12] с праймерами, фланкирующими маркерные участки аллелей S2 и S10 гена самонесовместимости [4]. Использовали следующие концентрации компонентов реакционной среды: В состав ПЦР смеси входили: 40-50 нг ДНК, 0,05мМ dNTPs, 0,3мкM каждого праймера, 1 единица активности (е. а.) Taq-ДНК полимеразы, в общем объеме реакционной смеси 25мкл. Постановку ПЦР осуществляли по следующей программе: 5 минут при 94о С - начальная денатурация; следующие 35 циклов: 30 секунд денатурация при 94о С, 40 секунд отжиг праймеров при 60о С, 30 секунд элонгация при 72о С; последний цикл элонгации 5 минут при 72о С.
Электрофорез продуктов ПЦР проводили в 2% агарозном геле, на основе трис боратного буфера при напряжении 150 V в течение 30 минут. Гелевые пластины окрашивали бромистым этидием и визуализировали в ультрафиолете.
Результаты
Получены результаты молекулярно-генетической идентификации целевых аллелей гена самонесовместимости для 27 кребов и 2 элитных селекционных форм яблони. В изученной выборке образцов идентифицировали обе целевые аллели. На рисунках 1 и 2 представлены результаты ПЦР идентификации данных аллелей у изучаемых образцов.
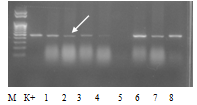
Рисунок 1 - Идентификация аллели S2 у образцов яблони
Примечания: М-маркер молекулярной массы ДНК; К+ - сорт контроль Гала; 1-8- исследуемые образцы яблони: 1- Спартак,
- 2 - Китайка № 3, 3 - 2-66-10, 4 - Транс Люценс, 5 - Империал Павла, 6 - Пиотош, 7 - Эксцельзиор, 8 - Желтогибридное
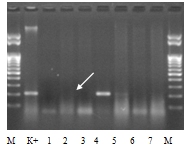
Рисунок 2 - Идентификация аллели S10 у образцов яблони
Примечания: М-маркер молекулярной массы ДНК; К+ - сорт контроль Мекинтош; 1-8 - исследуемые образцы яблони: 1- Желтое румяное, 2- Темно-вишневое, 3- Креб 2-71, 4- 12/1-20-34, 5- 12/2-20(24-28), 6- Геспер Роз, 7- Фейри
Выявление фрагмента на электрофореграмме на уровне, отмеченном стрелкой, соответствующем стандартному образцу, свидетельствует о наличии искомого аллеля гена самонесовместимости, отсутствие фрагмента - об отсутствии аллеля. В ходе исследования идентифицировали все целевые аллели, однако, с разной частотой встречаемости. Наиболее редким оказался аллель S10, который был выявлен у одного образца - элитной формы 12/1-20-34. Результаты генотипирования изученной выборки генотипов яблони по локусу S-гена представлены в таблице 1.
Таблица 1 - Идентификация аллелей S-гена у образцов яблони
|
№ п/п |
Сорт | ||
|
S2 |
S10 | ||
|
1 |
Пестрокрасное |
- |
- |
|
2 |
Красно-вишневое |
- |
- |
|
3 |
Краснополосатое |
- |
- |
|
4 |
Креб 67-2 |
+ |
- |
|
5 |
Китайка малиновая |
- |
- |
|
6 |
Рислинг красный |
- |
- |
|
7 |
Виктория |
- |
- |
|
8 |
Кетни |
- |
- |
|
9 |
Джон Дауни |
- |
- |
|
10 |
Долго |
+ |
- |
|
11 |
Спартак |
+ |
- |
|
12 |
Китайка № 3 |
+ |
- |
|
13 |
Креб 2-66-10 |
+ |
- |
|
14 |
Транс Люценс |
+ |
- |
|
15 |
Империал Павла |
- |
- |
|
16 |
Пиотош |
+ |
- |
|
17 |
Эксцельзиор |
+ |
- |
|
18 |
Желтогибридное |
+ |
- |
|
19 |
Желтое румяное |
+ |
- |
|
20 |
Темно-вишневое |
+ |
- |
|
21 |
Креб 2-71 |
+ |
- |
|
22 |
12/1-20-34 |
- |
+ |
|
23 |
12/2-20(24-28) |
+ |
- |
|
24 |
Геспер Роз |
+ |
- |
|
25 |
Фейри |
- |
- |
|
26 |
Желто-зеленое |
- |
- |
|
27 |
Никита |
- |
- |
|
28 |
Гертруда |
- |
- |
|
29 |
Экселенц Тиль |
- |
- |
Как видно из таблицы аллели идентифицируются со значительной разницей в частоте встречаемости. Если аллель S2 идентифицировался у 14 образцов (13 кребов и 1 элитной селекционной формы), то аллель S10 - лишь у одного образца (элитной формы 12/2-20(24-28). Примечательно, что аллель S10, относится к наиболее распространенным в мировом генофонде яблони, однако в изученной выборке образцов яблони он не распространен. Видно, что у кребов не идентифицируются одновременно два аллеля из представленных. В случае одновременного наличия у них двух изученных аллелей, которые являются наиболее распространенными, существенно возросла бы вероятность совпадения аллельного набора у креба и того промышленного сорта, для которого он предлагается как опылитель. На данном этапе работы, можно сделать заключение о том, что кребы Пестрокрасное, Красно-вишневое, Краснополосатое, Китайка малиновая, Рислинг красный, Виктория, Кетни, Джон Дауни, Империал Павла, Фейри, Желтозеленое, Никита, Гертруда и Экселенц Тиль можно рекомендовать, как более перспективные опылители из изученной выборки, так как у них не были выявлены аллели S2 и S10, которые относятся к числу наиболее распространенных в мировом генофонде яблони. Однако, необходимо сказать также и о целесообразности дальнейшего проведения молекулярно-генетической идентификации таких распространенных аллелей как S3, S5, S7, S1. Это даст возможность более обоснованно дать рекомендации по использованию изученных кребов в качестве опылителей для сортов, наиболее распространенных в промышленном садоводстве.
Литература
- 1 Хавкин Э. Е. Молекулярные маркеры в растениеводстве / Э. Е. Хавкин // Сельскохозяйственная биология. - 1997.- № 5.- С.3-19. 2 Суриков И. М. Несовместимость и эмбриональная стерильность растений. М. 1991. - 220 с. 3 Broothaerts W. cDNA cloning and molecular analysis of two self-incompatibility alleles from apple / W. Broothaerts, G. Janssens, P. Proost et al. // Plant Mol Biol. - V. 27. - 1995. -- P. 449-511. 4 Janssens G. A. A molecular method for S-allele identification in apple based on allele-specific PCR / G. A. Janssens, I. J. Goderis, W. F. Broekaert et al. // Theor. Appl. Genet. - 1995. - V. 91. - P. 691-698. 5 Broothaerts W. New findings in apple S-genotype analysis resolve previous confusion and request the re-numbering of some S-alleles // Theor Appl Genet. - V.106. _ 2003. - P. 703-714 6 Hegedыs A. Review of the self-incompatibility in apple (MalusЧdomestica Borkh., syn.: Malus pumila Mill.) // International Journal of Horticultural Science. V. 12. - 2006. - P. 31-36. 7 Супрун И. И. Молекулярно-генетическая идентификация аллелей гена самонесовместимости у сортов яблони отечественной селекции / И. И. Супрун, Е. В. Ульяновская, Я. В. Ушакова, Е. Т. Ильницкая // Доклады РАСХН. - 2011. - №5. - С. 15-17. 8 Супрун И. И. Использование методов ДНК-маркирования для идентификации аллелей гена самонесовместимости яблони // И. И. Супрун, Е. В. Ульяновская, Я. В. Ушакова / Труды Кубанского государственного университета. - №1(22), 2010. - с. 57-59. 9 Супрун И. И. Цитолого-генетическое изучение особенностей опыления иммунных и высокоустойчивых к парше сортов яблони // И. И. Супрун, Е. В. Ульяновская, Я. В. Ушакова / Агро ХХI. - №1-3, 2010. - с. 27-28. 10 Супрун И. И., Ульяновская Е. В. Изучение аллельного полиморфизма гена самонесовместимости и цитологических особенностей опыления сортов яблони // Плодоводство и виноградарство Юга России [Электронный ресурс] 2012. № 13 (01). URL: http://journal. kubansad. ru/pdf/12/01/02.pdf. 11 Murray, M. G. Rapid isolation of high molecular weight plant DNA / M. G. Murray, W. F. Thompson // Nucleic Acids Research, 1980.-V. 10.- Р. 4321-4325. 12 Шибата, Д. К. Полимеразная цепная реакция и молекулярно-генетический анализ биоптатов // Молекулярная клиническая диагностика. - М.: Мир, 1999. - С. 395-427.
Похожие статьи
-
Отбор и оценка селекционного материала Урожай зерна с единицы площади посева, наряду с продуктивностью зерна растения, находится в тесной зависимости от...
-
Потребу садивного матеріалу розраховують по кожному кварталу, потім вираховують загальну потребу по сортах на різних підщепах з урахуванням страхового...
-
Организационно-производственная структура хозяйства - это сочетание внутрихозяйственных производственных подразделений и аппарата управления, которое...
-
Выборка быков - производителей голштинской породы черно - пестрой масти включала 100 животных, рожденных в течение 23 лет в период с 1986 по 2009 год и...
-
Резервом збільшення виробництва насіння гетерозисних гібридів кукурудзи з одиниці площі необхідно забезпечити якнайповніше запилення качанів материнської...
-
Селекционная инвентаризация насаждений - Лесная селекция
Площадь отобранных насаждений должна составлять 20 30 % от всей площади насаждений, подвергшихся инвентаризации. Таблица 1 Ведомость плюсовых насаждений...
-
Общие рекомендации по проведении инсулинотерапии у кошек - Сахарный диабет у собак и кошек
Необходимо убедиться в том, что пациенту действительно требуется введение инсулина - у многих кошек может развиваться гипергликемия вследствие...
-
Продуктивное долголетие коров - Селекционные достижения в молочном скотоводстве
В условиях интенсификации производства молока продуктивное долголетие коров становится одним из основных критериев оценки их по пригодности к условиям...
-
Сроки, способы, формы внесения удобрений - Система удобрения в севообороте
Различают 3 способа внесения минеральных удобрений: основное, припосевное и подкормки. Основное удобрение. Предназначено для удовлетворения потребности...
-
Лесной питомник, Севообороты (Форма 2) - Лесные культуры
Лесной питомник - это предприятие, предназначенное для выращивания посадочного материала. Различают питомники по характеру выращиваемого посадочного...
-
Как было отмечено выше, ген BoLA-DRB3 обладает выраженным естественным полиморфизмом. Локус считается полиморфным, если в популяции существуют два или...
-
Синцитиальный метод при ранней идентификации вируса лейкоза крупного рогатого скота
Лейкоз крупного рогатого скота - злокачественное вирусное лимфопролиферативное заболевание. Ретровирус, относящийся к семейству Retroviridae, роду...
-
Клинические симптомы, как правило, неспецифичны и зависят от степени поражения внутренних органов и степени распространения возбудителя в организме...
-
В данном курсовом проекте для выполнения основных работ на лесосеке применяются укрупненные комплексные бригады (УКБ) и функциональные бригады (звенья)....
-
Основные принципы организации постоянной лесосеменной базы на селекционной основе - Лесная селекция
Семенной базой в лесном хозяйстве являются отобранные высокопроизводительные естественные насаждения и лесные культуры, а также специально сформированные...
-
Установление санитарно-защитных зон объектов - Установление санитарно-защитных зон объектов
На сегодняшний день болезненно актуальными для России продолжают оставаться вопросы, связанные с установлением санитарно-защитных зон промышленных...
-
Проблема коррекции агрессивного поведения собак
Вопросы агрессии занимают одно из ведущих направлений изучения специалистов, занимающихся поведением животных. Мы не знаем полных механизмов...
-
Выход чистой шерсти и его определение - Основы технологии животноводства
Выходом чистого волокна называют выраженное в процентах отношение массы чистого волокна при кондиционной влажности, с учетом остаточных компонентов, к...
-
1. Районирование сортов повышенной устойчивости. Наиболее восприимчивы к поражению фузариозом сорта твердых пшениц, а также сорта, характеризующиеся...
-
Первое упоминание о жимолости относится к XVII в., когда Камчатка была открыта В. Атласовым. Русские первопроходцы обратили внимание на дикорастущий...
-
Облепиха - удивительное растение, ежегодно дающее в условиях Сибири целебные плоды с прекрасным вкусом как в свежем, так и в переработанном виде. Плоды...
-
Устойчивость растений к патогенным микроорганизмам - Устойчивость растений
Жизнедеятельность растений нередко находится в сложных взаимосвязях с патогенными микроорганизмами и насекомыми. Способность противостоять заболеванию...
-
Идея и концепция решения проблемы (задача 1) Идея решения проблемы состоит в применении для этой цели современных IT-технологий, которых просто не...
-
Морфологическая характеристика - Подработка и сушка зерна риса
Морфология -- это учение о внешнем строении зерна и семян: форме, топографическом (пространственном) расположении частей зерна, размерах зерна и семени,...
-
Результаты гибридизации фейхоа (Feijoa Sellowiana Berg) во влажных субтропиках Краснодарского края
Результаты гибридизации фейхоа (Feijoa Sellowiana Berg) во влажных субтропиках Краснодарского края Субтропическая зона Краснодарского края является самой...
-
Агротехника овощных культур в открытом грунте. - Оценка хозяйства ОАО СельпромМаш
Арбуз. Все сорта арбуза относятся к виду, к одной ботанической разновидности - арбуз обыкновенный, столовый (). В диком виде растете на Африканском...
-
Биологические особенности картофеля Картофель (Solanum tuberosum) - это однолетнее травянистое растение. вырастающее до 100 сантиметров в высоту и...
-
Хозяйства для выращивания товарного картофеля должны использовать семенной материал не ниже 3-5 репродукции, поэтому оптимальным сроком обновления...
-
Морфологические признаки почв - Принципы растениеводства
Почва удобрение зерно Морфологические признаки - это внешние признаки почв. Для их исследования не нужно специальной лабораторной базы. Они изучаются...
-
Залежно від вибраної спеціалізації господарства доцільно вирощувати не більше 4-6 рекомендованих технічних і 3-4 столових сортів винограду, включених до...
-
Сортовой или гибридный состав - Черная смородина: посадка, выраивание, уход
Сегодня выведено множество сортов черной смородины, которые позволяют выбрать наиболее подходящий для любого региона нашей страны. Каждый сорт смородины...
-
Азотные удобрения - Луговое и полевое кормопроизводство
Основную долю в балансе почвенного азота составляют органические формы, находящиеся в гумусе, но они практически недоступны растениям без предварительной...
-
Обоснование вопроса Бурное развитие сельского хозяйства привели к увеличению продуктивности зернового хозяйства и скоплению на токах крупных колхозов и...
-
Пшеница Спельта(T. Spelta L.) - гр. Рубровелютинум. Обладает рядом полезных признаков: неприхотливость, сравнительная зимостойкость в ареале,...
-
IPO - Международный эквивалент Schutzhund - SchH (шутцхунд) с цифрой в качестве суффикса (1, 2, 3), который означает рабочую степень различной сложности....
-
Программирование урожая с.-х. культур - разработка комплекса технологических приемов, обеспечивающего оптимизацию регулируемых факторов среды для...
-
Согласно данным научной литературы на сегодняшний день существует множество теорий камнеобразования, в основе которых в качестве первопричины авторы...
-
Ботанічна характеристика та біологічні особливості кукурудзи Ботанічна характеристика кукурудзи У світовому рільництві, у тому числі й в Україні,...
-
Селекция и племенное дело в молочном скотоводстве - Селекционные достижения в молочном скотоводстве
Высокие темпы развития современного животноводства в развитых странах свидетельствуют о больших потенциальных возможностях селекции. На основе достижений...
-
Кормление молочных коров - Селекционные достижения в молочном скотоводстве
Кормление сельскохозяйственных животных является одним из определяющих факторов повышения продуктивности животных. При этом следует учесть, что ведущее...
Молекулярно-генетическая идентификация аллелей S2 и S10 гена самонесовместимости у кребов и элитных селекционных форм яблони