Гистолого-анатомические элементы прочности стебля овса посевного С. В. Лазаревич, С. П. Халецкий, З. В. Шемпель, С. С. Лазаревич
ВВЕДЕНИЕ
Главной методологической проблемой селекции овса посевного, как и других зерновых культур, является сочетание в одном генотипе признаков высокой продуктивности растений, качества продукции и устойчивости к неблагоприятным факторам внешней среды. Испытанным приемом создания таких генотипов является отбор по совокупности морфологических, технологических и биохимических признаков в полиморфных популяциях. Для оптимизации селекционных отборов на продуктивность злаков и устойчивость их к полеганию перспективным направлением представляется использование признаков анатомического строения стебля. Обширные исследования в этой области были выполнены на пшенице К. Г. Тетерятченко [1], В. В. Пыльневым [2] и другими исследователями. Анатомия стебля овса изучена в меньшей мере. Вместе с тем проблема устойчивости к полеганию является актуальной как для высокорослых, так и для короткостебельных сортов овса.
Целью наших исследований было выявление гистолого-анатомических элементов прочности стебля овса для оптимизации методов отбора. Задачами данной работы явилось изучение особенностей развития механических тканей стебля у короткостебельных сортообразцов, а также конструктивных особенностей проводящих пучков, влияющих на прочность соломины, которые могут быть рекомендованы для использования в селекции овса.
Анализ источников
Важным приспособлением высших растений к обитанию в наземных условиях является развитие у них комплекса тканей и анатомических структур, обеспечивающих прочность вегетативных органов. Прочность стебля зерновых культур обеспечивает устойчивость растений к динамическим и статическим деформирующим нагрузкам и в итоге - устойчивость к полеганию. В связи с этим изучению элементов прочности стебля уделяется большое внимание в селекции [3, 4], морфологии [5, 6, 7] и физиологии растений [8, 9].
Прочность является комплексным признаком. Она достигается сочетанием таких физических свойств, как упругость и жесткость. Упругость обеспечивает в пределах нормы реакции восстановление размеров и формы стебля после снятия деформирующей нагрузки, а жесткость характеризует способность противостоять механическим нагрузкам [10].
В результате исследований в полиплоидном ряду пшеницы установлено, что значения модуля упругости (Е) стебля существенно изменяются в зависимости от вида растения, изучаемого междоузлия и стадии развития растений [5]. Так, виды злаков с признаками ксероморфности имеют более жесткий стебель по сравнению с мезофитами; верхнее подколосовое междоузлие у них обладает большей упругостью, а нижнее - первое надземное, большей жесткостью; по мере созревания зерна прочность подколосового междоузлия увеличивается.
Проявление прочности стебля у злаков зависит от биохимического состава клеточных оболочек [8]. В частности, М. И. Лясковским установлено, что полегаемость посевов пшеницы в период колошение-налив зерна повышается в результате снижения содержания в стеблях целлюлозы, гемицеллюлозы и пектиновых веществ, которые используются растениями для синтеза олигосахаридов. Физические свойства стебля зависят также от ориентации микрофибрилл целлюлозы в клеточных стенках [11].
Большое влияние на прочность стебля оказывают его гистолого-анатомические структуры [5, 12], степень развития которых генетически детерминирована и может быть учтена в селекции на продуктивность и устойчивость к полеганию.
Методы исследования
Объектами исследований служили гистолого-анатомические признаки, обеспечивающие прочность стебля у короткостебельных образцов овса: Вандроунiк, Запавет, AV 17/3/10, Bandiccot, СI 8448, Coky, Flamingskurz, От - 207, Pennline, STH 815, 76Q : 28, 76Q : 225, (Вандроунiк Ч 76Q : 225) Ч Вандроунiк, (Белорусский голозерный Ч 76Q : 225) Ч Белорусский голозерный.
Растения выращивались на опытном поле лаборатории овса РУП "Научно-практический центр НАН Беларуси по земледелию". Почва опытного участка дерново-подзолистая, легкосуглинистая, развивающаяся на средних супесях, подстилаемых с глубины 0,7 м суглинистой мореной. Агрохимическая характеристика пахотного горизонта: содержание общего гумуса по Тюрину 1,9-2,2 %, рН в КСl - 6,0-6,2, подвижного фосфора (по Кирсанову) 225-239 и общего калия (по Масловой) 200-236 мг/кг почвы.
Отбор растений и фиксацию материала проводили в двухкратной повторности в начале выметывания метелки по общепринятым методикам цитологических исследований [13].
Микроскопические исследования проводились в УО БГСХА. Микропрепараты изготавливали из средних частей междоузлий, что позволило получить сопоставимые результаты. Для контрастирования на препаратах механических тканей и проводящих пучков срезы окрашивали флороглюцином. Верхнее, подметелочное, междоузлие обозначалось как EN1, второе сверху - EN2, третье - EN3, четвертое междоузлие - EN4.
Изучение микропрепаратов проводили с использованием оптического микроскопа Nikon Eclipse 50i, видеокамеры Nikon DS-Fi1, преобразователя сигналов Nikon digital sight и компьютера. Каждый линейный признак измерялся на каждом микропрепарате в пятикратном повторении. Статистическая обработка полученных результатов выполнена методом дисперсионного анализа [14] с использованием программы Microsoft Office Excel 2003.
ОСНОВНАЯ ЧАСТЬ
I. Общая характеристика гистолого-анатомических элементов прочности стебля овса посевного. Механическая прочность стебля овса, как и других мятликовых, обеспечивается несколькими гистолого-анатомическими факторами: наличием периферического кольца склеренхимы, а также проводящих пучков ПП пк (малые пучки, обслуживающие хлоренхиму первичной коры) и ПП пар. (большие пучки, расположенные в паренхиме центрального цилиндра); специфическим взаимным расположением проводящих пучков; лигнификацией клеток паренхимы центрального цилиндра, находящихся между проводящими пучками вблизи кольца склеренхимы.
Периферическое кольцо склеренхимы (рис.1, а) образуется в результате дифференциации клеток перицикла. Оно располагается на периферии стебля под эпидермисом, окружая попарно расположенные тяжи хлоренхимы (рис.1, з) и находящиеся между ними малые проводящие пучки (ПП пк) (рис.1, б). Прозенхимные клетки этой склеренхимы вытянуты вдоль оси стебля, имеют малый диаметр поперечного сечения и плотное сложение. Их толстые оболочки пропитаны лигнином. Этой ткани принадлежит ведущая роль в формировании упругости стебля.

Рис. 1 - Анатомическое строение стебля овса в подметелочном междоузлии: а - склеренхима перициклического происхождения; б - проводящий пучок ПП пк; в - склеренхимная обкладка пучка ПП пк; г - проводящий пучок ПП пар.; д - склеренхимная обкладка пучка ПП пар.; е - тяж склеренхимы между сосудами ПП пар.; ж - паренхима центрального цилиндра; з - хлоренхима
Конструкция проводящих пучков также обеспечивает прочность стебля, тем более, что они окружены склеренхимной обкладкой (рис.1, в, д), возникшей в результате деятельности клеток прокамбия. Эта обкладка лучше развита на радиально противоположных сторонах пучков (2-5 рядов клеток), подвергающихся большим деформирующим нагрузкам, возникающим при изгибании стебля. Тангентальные стороны обкладки испытывают меньшие физические нагрузки и образованы лишь одним рядом клеток склеренхимы.
Между сосудами метаксилемы пучков обоих типов обнаруживаются 2-4 ряда клеток мелкоклеточной склеренхимы (рис.1, е), которые повышают прочность строения проводящих пучков. В процессе формирования стебля эта ткань образуется одновременно с появлением сосудов метаксилемы. На более ранних этапах образования проводящих пучков при наличии в составе ксилемы только сосудов протоксилемы эта склеренхимная ткань отсутствует.
Проводящие пучки ПП пк и ПП пар., находящиеся на разном удалении от поверхности стебля, гармонично чередуются между собой так, что их геометрические центры представляют собой вершины треугольников (рис.1, 2, 3, 4). Такое расположение пучков повышает жесткость анатомической конструкции стебля. Оно имеет особое значение в подметелочном междоузлии, где препятствует пониканию метелок.
Из других анатомических приспособлений, повышающих прочность соломины, можно отметить ребристость поверхности стебля (рис. 2), обнаруженную в верхнем междоузлии у некоторых образцов овса.

Рис. 2 - Ребристость поверхности стебля овса: а - склеренхима перициклического происхождения; б - проводящий пучок ПП пар.; в - проводящий пучок ПП пк; г - выступ поверхности стебля
Поверхностные выступы образуются в местах расположения проводящих пучков ПП пк. В этом случае склеренхима перициклического происхождения на участках между выступами (ребрами) развита слабо и состоит лишь из 3-4 слоев клеток. Однако при этом каждый сегмент поверхностного слоя склеренхимы имеет вид швеллера, жесткость которого выше, чем у гладкой полосы. Такой признак может иметь значимость для образцов с тонкой стенкой соломины.
В междоузлиях средней и нижней частей стебля овса в толще клеток паренхимы центрального цилиндра иногда встречаются сплошные тяжи механических тканей (рис. 3). Исследования, проведенные на разных видах пшеницы, показали, что такие тяжи являются следствием реконструкции проводящих пучков в узлах побегов [5].
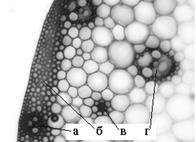
Рис. 3 - Тяж механических тканей в стебле овса: а - проводящий пучок ПП пк; б - склеренхима перициклического происхождения; в - тяж склеренхимы; г - проводящий пучок ПП пар
Они изредка встречаются в междоузлиях средней части стебля и не выходят в очередное, вышерасположенное, междоузлие.
Уникальная конструкция стебля была обнаружена у короткостебельного низкопродуктивного образца AV 17/3/10 (рис. 4). В его междоузлиях анатомические структуры, обеспечивающие прочность стебля оказались неразвитыми.
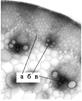
Рис. 4 - Поперечный срез стебля образца AV 17/3/10: а - полоса неразвитой склеренхимы перициклического происхождения; б - проводящий пучок ПП пк; в - проводящий пучок ПП пар
Во-первых, клетки перицикла (рис.4, а) не были дифференцированы в типичную склеренхиму. Их клеточные оболочки оставались тонкими, а обработка среза флороглюцином выявила лишь следы лигнина в них. Во-вторых, отсутствовала склеренхимная обкладка проводящих пучков ПП пк и ПП пар. В-третьих, в пучках ПП пар. между сосудами метаксилемы мелкоклеточная склеренхима образует лишь один ряд клеток. Вместе с тем стенки сосудов были типично насыщены лигнином. Отмеченные особенности строения стебля явились первопричиной сильной полегаемости растений короткостебельного образца AV 17/3/10 в полевых условиях.
II. Влияние генетических особенностей на развитие количественных гистолого-анатомических признаков стебля, обеспечивающих устойчивость овса посевного к полеганию.
Развитие гистолого-анатомических признаков, оказывающих влияние на прочность стебля у одновозрастных растений овса, зависит от генотипа изученного образца и места нахождения междоузлия в системе целостного побега. Об этом свидетельствует значительное превышение фактических значений критерия Фишера над теоретически ожидаемыми на 5 %-ном уровне значимости (табл. 1).
Таблица 1 - Результаты дисперсионного анализа признаков стебля сортообразцов овса посевного
|
Признак стебля |
Фактор варьирования признака |
F фактич. |
F05 |
|
Толщина соломины |
Сорт Междоузлие |
|
|
|
Толщина стенки соломины |
Сорт Междоузлие |
|
|
|
Число проводящих пучков ПП пк |
Сорт Междоузлие |
|
|
|
Число проводящих пучков ПП пар. |
Сорт Междоузлие |
|
|
|
Толщина периферического кольца склеренхимы перициклического происхождения |
Сорт Междоузлие |
|
|
|
Число рядов клеток в периферическом кольце склеренхимы перициклического происхождения |
Сорт Междоузлие |
|
|
Для объяснения причин разной устойчивости изученных короткостебельных сортообразцов овса к полеганию при микроскопическом анализе учитывались гистолого-анатомические признаки, обеспечивающие прочность стебля в разных междоузлиях. Результаты исследований показали, что у всех изученных сортообразцов толщина слоя склеренхимы перициклического происхождения, а также число рядов клеток в слое склеренхимы и диаметр клеток этой ткани закономерно изменяются при перемещении от нижнего к подметелочному междоузлию (табл. 2). Так, толщина периферического слоя склеренхимы изменяется пропорционально изменению толщины стенки соломины. Относительно широкий слой склеренхимы в нижнем междоузлии (EN4) уменьшается в середине стебля (EN3 и EN2), а затем снова увеличивается в подметелочном междоузлии (EN1). Причем у большинства сортообразцов наиболее широкий слой склеренхимы формируется в подметелочном междоузлии. Прирост толщины слоя склеренхимы в EN1 по сравнению с EN2 превышал 30 % [(Белорусский голозерный Ч 76Q : 225) Ч Белорусский голозерный, СI 8448, AV 17/3/10]. Это увеличение толщины слоя механической ткани предохраняет растения от поникания и обламывания метелок.
Таблица 2 - Развитие периферического кольца склеренхимы в стебле овса
|
Сортообразец |
Толщина периферического слоя склеренхимы, мкм |
Число рядов клеток в периферическом слое склеренхимы, шт. |
Толщина клеток Склеренхимы, мкм | |||||||
|
В EN 1 |
В EN 2 |
В EN 3 |
В EN 4 |
В EN 1 |
В EN 2 |
В EN 3 |
В EN 1 |
В EN 2 |
В EN 3 | |
|
Запавет |
64,8 |
50,1 |
51,6 |
62,9 |
5,2 |
3,8 |
3,6 |
12,5 |
13,2 |
14,3 |
|
76Q : 225 |
96,1 |
70,8 |
67,1 |
80,8 |
6,8 |
4,2 |
4,4 |
14,1 |
16,9 |
15,2 |
|
76Q : 28 |
77,0 |
67,2 |
65,0 |
78,9 |
6,2 |
4,3 |
4,4 |
12,4 |
15,6 |
14,8 |
|
Coky |
72,8 |
61,0 |
51,4 |
60,0 |
5,6 |
3,9 |
3,5 |
13,0 |
15,6 |
14,7 |
|
СI 8448 |
84,1 |
61,4 |
58,2 |
71,3 |
6,7 |
4,9 |
4,0 |
12,6 |
12,5 |
14,6 |
|
От - 207 |
79,1 |
68,9 |
65,6 |
74,8 |
5,7 |
4,3 |
4,2 |
13,9 |
16,0 |
15,6 |
|
Pennline |
92,0 |
93,4 |
79,3 |
95,0 |
5,9 |
4,9 |
4,7 |
15,6 |
19,1 |
16,9 |
|
AV 17/3/10 |
138,2 |
101,7 |
124,2 |
141,2 |
8,2 |
5,9 |
6,1 |
16,9 |
17,2 |
20,4 |
|
Flamingskurz |
81,2 |
74,5 |
65,0 |
67,1 |
6,1 |
4,4 |
4,5 |
13,3 |
16,9 |
14,4 |
|
STH 815 |
89,2 |
67,0 |
69,6 |
70,7 |
7,1 |
4,4 |
4,2 |
12,6 |
15,2 |
16,6 |
|
Вандроунiк |
62,5 |
54,4 |
57,9 |
54,0 |
4,7 |
3,9 |
3,8 |
13,3 |
13,9 |
15,2 |
|
(Бел. голозерный Ч 76Q : 225) Ч Бел. голозерный |
104,7 |
65,9 |
57,6 |
61,4 |
7,0 |
4,4 |
3,9 |
15,0 |
15,0 |
14,8 |
|
(Вандроунiк Ч 76Q : 225) Ч Вандроунiк |
92,8 |
68,6 |
61,0 |
69,8 |
7,1 |
4,4 |
3,9 |
13,1 |
15,5 |
15,6 |
|
Bandiccot |
94,6 |
62,0 |
56,8 |
101,2 |
7,2 |
4,0 |
3,7 |
13,1 |
15,5 |
15,4 |
|
НСР05 по фактору "сорт" |
8,6 |
0,8 | ||||||||
|
НСР05 по фактору "междоузлие" |
3,3 |
0,3 |
У всех изученных растений периферическое кольцо склеренхимы в подметелочном междоузлии отличалось малым диаметром клеток на поперечном срезе и увеличенным количеством их слоев. Вместе с тем образцы овса существенно различались по этим признакам. Например, сорт Запавет, обладающий признаками ксероморфности по развитию проводящих тканей, отличался малым диаметром клеток склеренхимы в EN1 (12,5 мкм) и небольшим числом их рядов (5,2). Более крупные клетки диаметром 15-16 мкм были характерны для образцов (Белорусский голозерный Ч 76Q : 225) Ч Белорусский голозерный, Pennline, AV 17/3/10.
Для формирования устойчивости растений овса к полеганию большое значение имеет толщина клеточных оболочек склеренхимы и инкрустированность их лигнином. Визуально более толстые клеточные оболочки склеренхимы, хорошо окрашиваемые флороглюцином, были характерны для устойчивых к полеганию образцов 76Q : 225, Flamingskurz и Bandiccot (табл. 3). Степень полегания этих растений составила лишь 1 балл. Тонкостенные клетки склеренхимы с крайне низким уровнем лигнификации были обнаружены у полегающего образца AV 17/3/10 (степень полегания - 9 баллов).
Таблица 3 - Морфолого-анатомическая характеристика короткостебельных сортообразцов овса
|
Сортообразец |
Высота растений, См |
Диаметр Стебля, Мм |
Подметелочное междоузлие |
Степень полегания перед уборкой, балл | |||
|
Выполненность стебля, % |
ИТС, % |
Число ПП пар., Шт. |
Число ПП пк, Шт. | ||||
|
Запавет |
105,8 |
3,3 |
53,8 |
12,2 |
23,8 |
22,5 |
5 |
|
76Q : 225 |
61,0 |
2,8 |
65,0 |
16,8 |
13,6 |
14,9 |
1 |
|
76Q : 28 |
65,2 |
2,9 |
63,5 |
13,4 |
14,3 |
15,0 |
3 |
|
Coky |
73,2 |
2,5 |
57,8 |
16,6 |
15,1 |
16,4 |
1 |
|
СI 8448 |
54,8 |
3,0 |
68,8 |
12,7 |
18,9 |
18,6 |
5 |
|
От - 207 |
62,4 |
2,9 |
70,1 |
12,0 |
14,3 |
16,5 |
1 |
|
Pennline |
83,0 |
3,7 |
60,9 |
13,3 |
15,7 |
17,0 |
1 |
|
AV 17/3/10 |
39,4 |
2,7 |
74,1 |
20,8 |
10,7 |
11,6 |
9 |
|
Flamingskurz |
75,4 |
3,3 |
68,2 |
12,6 |
18,5 |
19,5 |
1 |
|
STH 815 |
65,8 |
2,9 |
72,4 |
13,0 |
15,9 |
15,9 |
7 |
|
Вандроунiк |
105,8 |
3,2 |
56,6 |
11,4 |
20,7 |
19,9 |
3 |
|
(Бел. голозерный Ч 76Q : 225) Ч Бел. голозерный |
68,4 |
3,2 |
75,0 |
13,1 |
20,3 |
21,6 |
1 |
|
(Вандроунiк Ч 76Q : 225) Ч Вандроунiк |
58,8 |
3,4 |
72,8 |
11,4 |
20,0 |
23,1 |
3 |
|
Bandiccot |
61,0 |
2,6 |
72,5 |
15,3 |
15,0 |
16,4 |
1 |
|
НСР05 |
0,3 |
1,4 |
1,4 |
Овес стебель устойчивость ткань
Развитие механических тканей в разных междоузлиях стебля овса контролируется единой генетической системой. Поэтому количественные признаки склеренхимы достаточно тесно коррелируют между собой. Так, коэффициент корреляции (r) толщины слоя склеренхимы в EN1 и EN2 в группе из 14 сортообразцов был равен 0,58, а для EN2 и EN3 - r = 0,63. В связи с этим для сравнительной оценки потенциальной прочности стебля изучаемых сортообразцов возможен учет признаков склеренхимы лишь в одном однотипном междоузлии. Это существенно уменьшит количество фиксируемых для анализа междоузлий, а также количество изучаемых микропрепаратов.
Выбор междоузлия может корректироваться в соответствии с конкретными задачами селекции. Например, анализ тканей в верхнем междоузлии (EN1) окажется рациональным при отборе генотипов с непоникающей и необламывающейся метелкой, а в EN2 или EN3 - при отборе образцов, устойчивых к стеблевому полеганию. Устойчивость сортообразцов овса к полеганию определялась сочетанием разных морфологических и анатомических признаков стебля (табл. 3).
Например, высокая устойчивость образца 76Q : 225 была связана с хорошей выполненностью [5] соломины (65 %) в подметелочном междоузлии и хорошим развитием периферического слоя склеренхимы, при котором отношение толщины слоя склеренхимы к толщине стенки соломины (ИТС) составило 16,8 %. Устойчивость образца Flamingskurz обеспечивалась толстой (3,3 мм), хорошо выполненной соломиной (68,2 %), большим числом проводящих пучков (18,5 ПП пар. и 19,5 ПП пк). У образца (Белорусский голозерный Ч 76Q : 225) Ч Белорусский голозерный избыточная выполненность стебля (75,0 %) была компенсирована большим числом проводящих пучков (20,3 ПП пар. и 21,6 ПП пк) и средним развитием периферического кольца склеренхимы (ИТС=13,1 %). Полегаемость растений сорта Запавет объясняется слабой выполненностью стебля (53,8 %); образца AV 17/3/10 - избыточной выполненностью стебля (74,1 %), небольшим числом проводящих пучков (10,7 ПП пар. и 11,6 ПП пк) при слабом развитии склеренхимы перициклического происхождения. Эти данные свидетельствуют о необходимости учета разных гистолого-анатомических признаков при оценке генотипов овса на устойчивость к полеганию.
ЗАКЛЮЧЕНИЕ
При проведении отборов среди низкорослых образцов овса на продуктивность и устойчивость к полеганию следует учитывать комплекс морфологических и гистолого-анатомических признаков и отдавать предпочтение толстостебельным растениям с выполненностью стебля около 61,8 %, с большим числом проводящих пучков среднего диаметра и широким полем флоэмы [15], с широким периферическим кольцом мелкоклеточной склеренхимы перициклического происхождения.
Анализ развития механических тканей следует проводить в однотипных междоузлиях.
ЛИТЕРАТУРА
- 1. Тетерятченко, К. Г. Анатомический метод оценки исходного материала мягкой озимой пшеницы на продуктивность, морозостойкость и устойчивость к полеганию / К. Г. Тетерятченко // Науч.-техн. бюл. / ВИР им. Н. И. Вавилова. - 1984. Вып. 146. - С. 28-32. 2. Пыльнев, В. В. Изменение анатомического строения растений озимой пшеницы в результате селекции / В. В. Пыльнев, Б. Б. Батоев // Известия ТСХА. - М.: 1993. - Вып. 1. - С. 31-39. 3. Ильинская-Центилович, М. А. Устойчивость к полеганию как проблема селекции озимой пшеницы: автореф. дис. ... д-ра с.- х. наук / М. А. Ильинская-Центилович; Харьковский с. - х. ин-т им. В. В. Докучаева. - Харьков, 1964. - 48 с. 4. Бороевич, С. Принципы и методы селекции растений / С. Бороевич / Пер. с сербохорв. В. В. Иноземцева; Под ред. и с предисл. А. К. Федорова. - М.: Колос, 1984. - 344 с. 5. Лазаревич, С. В. Эволюция анатомического строения стебля пшеницы / С. В. Лазаревич. - Минск: БИТ "Хата", 1999. - 296 с. 6. Дорофеев, В. Ф. Анатомическое строение стебля некоторых видов пшениц и его связь с полеганием / В. Ф. Дорофеев // Бот. журнал. - 1962. - № 47. - С. 377-380. 7. Мережко, А. Ф. Наследование длины соломины и анатомического строения стебля межсортовыми гибридами мягкой пшеницы / А. Ф. Мережко, О. Д. Градчанинова // Бюл. ВИР им. Н. И. Вавилова. - 1982. - Вып. 122. - С. 18-22. 8. Лясковский, М. И. Динамика фенольных соединений и лигнина в стебле озимой пшеницы и формирование устойчивости к полеганию / М. И. Лясковский, Ф. Л. Калинин // Физиология и биохимия культурных растений. - 1997. - Т.9, № 4. - С. 359-365. 9. Brozman, D. Determination of Young, s modulus of elasticity in tention of corn culm, by the speckle interferometry metod / D. Brozman // Acta Technol. Agricult. - 1992. - Vol.32. - P. 19-24. 10. Напряжения и деформации при растяжении и сжатии в пределах упругости. Подбор сечений / Беляев Н. М. Сопротивление материалов. - 15-е изд., перераб. - М.: Наука, 1976. - С. 26-38. 11. Zebrowski J. Lamliwosc zdzbla pszenicy w swietle ultrastrukturalnych badan scian komorkowych || Biuletyn Instytutu Hodowli i Aklimatyzacji Roslin. - 1989. № 171 - 172. - P. 187-189. 12. Гудкова, Г. Н. Сравнительно-анатомическое изучение вегетативных органов некоторых сортов ржи в связи с их различной устойчивостью к полеганию / Г. Н. Гудкова // Труды / Ленингр. о-во естествоиспыт. - 1972. Т. 72. - Вып. 3. - С. 34-42. 13. Паушева, З. П. Практикум по цитологии растений. - 4-е изд., перераб. и доп./ З. П. Паушева. - М.: Агропромиздат, 1988. - С. 61-66. 14. Доспехов, Б. А. Методика полевого опыта (с основами статистической обработки результатов исследований) / Б. А. Доспехов. - М.:Агропромиздат, 1985. - С. 207-213. 15. Лазаревич, С. В. Развитие проводящей системы у короткостебельных сортообразцов овса посевного / С. В. Лазаревич, С. П. Халецкий, А. Г. Власов, И. Г. Матыс, С. С. Лазаревич // Вестник Белорусской государственной сельскохозяйственной академии. - 2013. - № 2. - С. 66-71. УДК 635.8.032.034
Похожие статьи
-
Для того чтобы грамотно проводить подкормки растений в ходе вегетации необходимо проводить диагностику обеспеченности растений необходимыми питательными...
-
Роль отдельных химических элементов - Обмен минеральных веществ и его нарушения
Натрий и хлор. Эти элементы поступают в организм в виде хлорида натрия, в том же виде выводятся из него и связаны между собой в процессе обмена веществ....
-
Интенсивная технология возделывания овса - Технологические особенности возделывания овса
Место в севообороте. В отличие от других зерновых злаков овес слабо поражается корневыми гнилями, поэтому при достаточном внесении удобрений различия в...
-
Зерно овса содержит 13...14% белка, 5...6% жира, 40...45% крахмала. Белок овса содержит ценные незаменимые аминокислоты - лизин, триптофан. Зерно богато...
-
Значение основных элементов питания в жизни растений - Почвоведение как наука
Для правильного и спокойного роста растений и их развития из семян и до зрелости необходимо 16 различных элементов. Три из этих элементов - углерод,...
-
Устойчивость растений к патогенным микроорганизмам - Устойчивость растений
Жизнедеятельность растений нередко находится в сложных взаимосвязях с патогенными микроорганизмами и насекомыми. Способность противостоять заболеванию...
-
Калийные удобрения - Качественные свойства и признаки почв
Калийное удобрение является одним из самых необходимых элементов минерального питания растений. Калий не входит в состав органических соединений в...
-
Для нормального роста и развития растений необходимы различные элементы питания. По современным данным, таких элементов порядка 20, без которых растения...
-
Особенности посева (посадки) чечевицы на зерно в Радищевском районе Ульяновской области Сроки посева Способы посева, ширина междурядий, см Норма высева...
-
Таблица 12. Посевные качества семян Показатели Масса 1000 семян, г Энергия прораста-ния, % Всхо-жесть, % Семена с растений без чеканки 16 93,5 98,5...
-
Биологические особенности картофеля Картофель (Solanum tuberosum) - это однолетнее травянистое растение. вырастающее до 100 сантиметров в высоту и...
-
Жизненный цикл зерновых хлебов - Зерновые культуры Республики Беларусь. Их основные болезни
В течение вегетации зерновые культуры проходят ряд фенологических фаз, которые отличаются друг от друга появлением новых органов и рядом внешних...
-
Заключение - Сельская агрометеорология
Одной из главных причин снижения урожайности высокопродуктивных сельскохозяйственных растений является их недостаточная устойчивость к неблагоприятным...
-
Появление картофеля в России связывают с петровской эпохой. - Свойства картофеля
Один из основоположников научного картофелеводства в России агроном А. Т. Болотов, в 1770г опубликовал работу "Примечание о картофеле, или земляных...
-
Биологические особенности ржи. Рожь (Secale cereale) имеет более широкие листья, чем пшеница. Всходы у ржи коричнево-фиолетовой окраски. Колосковые...
-
Основное преимущество клонального микроразмножения - получение генетически однородного, безвирусного посадочного материала. Предположение о возможности...
-
Размер левад. Группа лошадей Левады Норма площади на 1 м Жеребцы Индивидуальные 600 Кобылы Групповые 20 Молодняк Групповые 20 В левадах, примыкающих к...
-
Введение Внедрение экологически безопасных, адаптивных технологий, в том числе культивирование смешанных посевов различных культур и сортов, является...
-
Обоснование выбора, описание устройства и работ Рычажное приспособление Состоит из зацепа 2, в который ввернуты рукоятка 1 и винт 3 с резьбой М 10. Чтобы...
-
Для оценки функциональной полноценности иммунной системы и диагностики ее нарушений необходимо применять клинические, патологоанато - мические и...
-
Уборка, послеуборочная обработка и реализация овса - Технология хранения продукции растениеводства
Зерно овса - прекрасный концентрированный корм. Оно имеет большое значение при выращивании молодняка и птицы, при откорме животных. В зерне овса...
-
Конечной целью возделывания конопли является получение продукции - семян, тресты. Получение ее в запланированном количестве возможно лишь при...
-
Очень важно было выяснить значение каждого из элементов слагаемых структуры урожая в условиях сухой степи в зависимости от возделывания по паровому и...
-
В составе здания основного назначения есть следующие помещения : А) помещение для телят Б) молочная В) моечная Г) помещение для хранения текущего запаса...
-
Чечевица в Ульяновской области новая зерновая бобовая культура, особенно переспективная для получения высокобелковых добавок в пищевых целях. Однако,...
-
При обмолоте зерно сразу очищают от сорняков и других примесей на ворохоочистительных машинах, при необходимости сушат до влажности 14-15%, затем...
-
Сложность механизации уборки чечевицы обусловливается ее биологическими особенностями -- осыпаемостью, низким расположением бобов. Основной способ уборки...
-
Чечевица, как и другие зернобобовые культуры, хорошо использует последействие органических и минеральных удобрений, отличается повышенным усвоением...
-
Размещение культуры в севообороте Важное значение для получения устойчивых урожаев имеет правильное размещение чечевицы в севообороте. На приусадебном...
-
Фазы роста и развития (на примере зерновых хлебов) Фаза Признаки данной фазы Практическое значение В выращивании и получении продукции 1. Всходы...
-
7. Химический состав и питательная ценность чечевицы Используемые части растения Содержание, % В 1 кг корма Вода протеин Жир Клетчатка БЭВ Зола К. ед...
-
Требование к почве и элементам питания - Агротехника гороха. Расчет величины урожая
Лучшими для гороха являются нейтральные среднетяжелые суглинистые и супесчаные плодородные почвы, содержащие в достаточном количестве перегной и известь...
-
Свойства почвы, обуславливающие необходимость известкования и гипсования - Почвоведение как наука
Известкование и гипсование почв - Известкование имеет двойное значение: известь улучшает физико-химические свойства почвы, создавая тем самым...
-
Техническое планирование, Технологический процесс - Теплица по выращиванию овощей (помидоры, огурцы)
Технологический процесс Ведение двух культур в несколько оборотов требует изолированного рассадного отделения для обеспечения независимого от основных...
-
Требование к теплу и свету, Требование к влаге - Агротехника гороха. Расчет величины урожая
Горох -- холодостойкое растение, может переносить заморозки до --5°С, молодые побеги и цветки -- до --2...--3°С. Оптимальная температура в период...
-
Уход за виноградниками - Организация работы виноградников
Виноградные лозы всегда тянутся вверх, поближе к солнцу. Если не следить за их ростом и пустить этот процесс на самотек, то растения будут обвивать...
-
Вывод и предложения - Агропромышленный комплекс республики: состояние и перспективы развития
Технология должна планироваться с учетом местных природно-климатических и организационно-технических условий, но при этом необходимо учитывать...
-
Истребительные мероприятия, Агротехнические - Защита сельскохозяйственных растений от болезней
Агротехнические Агротехнические (провокация, механическое и физическое уничтожение, истощение, удушение, высушивание, вымораживание и др.) Для...
-
Уход за посевами, Уборка урожая - Луговое и полевое кормопроизводство
Обязательный прием по уходу -- прикатывание почвы после посева. При большой глубине заделки семян после увлажнения верхнего слоя почвы необходимо...
-
Заключение, Список используемых источников - Клональное размножение и оздоровление растений
В целом методы клонального микроразмножения, несомненно, имеют ряд преимуществ перед существующими традиционными способами размножения: - получение...
Гистолого-анатомические элементы прочности стебля овса посевного С. В. Лазаревич, С. П. Халецкий, З. В. Шемпель, С. С. Лазаревич