Структурно-функціональні особливості міокарда теличок за різної рухової активності - Морфофункціональні особливості міокарда теличок за різної рухової активності
Особливістю робочих тканинних структур серця добових теличок є їхня незавершеність, що проявляється поздовжньою посмугованістю. У міокарді правого передсердя кардіоміоцити розташовуються окремими пучками, між якими розміщується пухка волокниста сполучна тканина. Кардіоміоцити у пучках щільно прилягають один до одного, мають оксіфильну саркоплазму, що містить міофібрили, ледве помітні на поперечних зрізах з поздовжньою посмугованістью, тоді як поперечна майже не виявляється. У правому передсерді провідні кардіоміоцити відсутні, а відносна площа робочих складає 88,15 % (табл. 1). Відносна площа кровоносних судин переважає в міокарді лівого передсердя. У міокарді правого шлуночка реєструються слабковиражені межі між кардіоміоцитами, що контактують "кінець у кінець". Товщина кардіоміоцитів правого шлуночка на 10,0 % більша, ніж правого, і на 33,61 % - лівого передсердь, а довжина їх ядер, відповідно, на 7,96 і 13,55 %. У лівому передсерді кардіоміоцити також розташовуються окремими пучками. Відносна площа робочих кардіоміоцитів на 3,2 % менше, ніж у правому передсерді. Товщина кардіоміоцитів у лівому передсерді на 20,78 % більша, ніж у правому, а довжина їх ядер, навпаки, на 5,17 % менше. Товщина кардіоміоцитів лівого і правого шлуночків практично однакова (4,760,18 і 4,730,28 мкм; табл. 1). Діаметр кардіоміоцитів у лівому передсерді добових теличок на 16,3 %, а їх об'єм - на 32,10 % менші, ніж у правому(табл. 2). Ядра кардіоміоцитів овальні, у багатьох виявлений гетерохроматин. Ядерно-цитоплазматичне відношення в два рази перевершує в міокарді правого передсердя.
Таблиця 1 - Морфомеричні показники міокарда серця добових теличок
|
Показник |
Передсердя |
Шлуночки |
Міжшлуночкова перегородка | ||
|
Праве |
Ліве |
Правий |
Лівий | ||
|
Відносна площа КМЦ*, % |
88,152,45 |
85,383,82 |
92,484,79 |
90,447,51 |
93,434,21 |
|
Площа робочих КМЦ, у проц. |
88,152,45 |
85,383,82 |
82,964,32 |
80,032,77 |
82,911,98 |
|
Площа провідних КМЦ, у проц. |
- |
- |
9,520,70 |
10,410,19 |
10,520,21 |
|
Товщина робочих КМЦ, мкм |
4,300,27 |
3,560,15 |
4,730,28 |
4,760,18 |
4,900,14 |
|
Довжина ядер робочих КМЦ, мкм |
8,540,36 |
8,120,25 |
9,220,26 |
8,790,42 |
6,200,17 |
|
Площа кровоносних Судин, у проц. |
6,350,30 |
9,660,45 |
4,600,20 |
6,940,68 |
3,800,14 |
|
Площа пухкої волокнистої сполучної тканини, у проц. |
5,500,20 |
4,960,76 |
2,920,18 |
2,620,24 |
2,770,08 |
* КМЦ - кардіоміоцити
У міокарді лівого передсердя площа поперечного перерізу ядер робочих кардіоміоцитів на 9,2, а цитоплазми 16,8 % менша, ніж правого. Діаметр кардіоміоцитів і ядер у міокарді правого шлуночка також перевищує показники міокарда передсердь. Об'єм ядер міокарда правого шлуночка на 33,7 % більший, ніж у правому, і на 96,92 %, ніж у лівому передсердях. У міокарді правого шлуночка площа поперечного перерізу ядер на 13,14 % більша, ніж у правому передсерді, і на 24,59 %, порівняно з лівим. Ядра кардіоміоцитів у правому шлуночку більш видовжені і їхній коефіцієнт складає 2,020,02 проти 1,950,03 у правому передсерді. У кардіоміоцитах лівого шлуночка чітко виявляються міофібрили, для яких характерна поздовжня орієнтація. Каріометричні структури міокарда лівого шлуночка превалюють, порівняно з правим (табл. 2).
У міокарді лівого шлуночка найменша відносна площа пухкої волокнистої сполучної тканини. Відносна площа кровоносних судин міокарда лівого шлуночка на 50,87 % більша, порівняно з правим. У міжшлуночковій перегородці відносна площа кардіоміоцитів найбільша. Товщина їх на 2,94 % зростає у лівому і на 3,59 % - правому шлуночках, порівняно з передсердями - на 13,95 і 37,64 %. Довжина ядер кардіоміоцитів у перегородці менша, ніж у лівому (32,76%) і правому (29,47%) шлуночках. Об'єм ядер у міжшлуночковій перегородці максимальний. Ядра більш видовжені і коефіцієнт складає 2,030,03. Відносна площа поперечного перерізу ядер кардіоміоцитів і цитоплазми також менша, ніж у шлуночках та передсердях.
Таблиця 2 - Морфометричні показники ядер робочих кардіоміоцитів міокарда серця добових теличок
|
Показник |
Передсердя |
Шлуночки |
Міжшлуночкова Перегородка | ||
|
Праве |
Ліве |
Правий |
Лівий | ||
|
Діаметр КМЦ, мкм |
7,850,09 |
6,570,08 |
8,110,08 |
9,420,21 |
8,270,09 |
|
Поперечний діаметр КМЦ, мкм |
3,880,05 |
3,330,05 |
4,390,07 |
5,290,12 |
4,180,03 |
|
V-об'єм ядра КМЦ, мкм3 |
79,913,85 |
54,252,27 |
106,833,24 |
112,343,56 |
121,122,94 |
|
Е-коефіцієнт подовженості ядра КМЦ |
1,950,03 |
2,960,05 |
2,020,02 |
1,770,02 |
2,030,03 |
|
Ядерно-цитоплазматичне відношення (N/P) |
0,240,004 |
0,630,002 |
0,540,007 |
0,560,007 |
0,570,07 |
|
S-площа поперечного перерізу ядра КМЦ, мкм2 |
6,090,34 |
5,530,26 |
6,890,15 |
8,300,19 |
6,570,05 |
|
S1- площа поперечного перерізу саркоплазми, КМЦ, мкм2 |
12,350,15 |
10,280,14 |
12,730,12 |
14,790,32 |
12,980,15 |
Ядерно-цитоплазматичне відношення особливої різниці не має. Пухка волокниста сполучна тканина складає у міокарді всього 2,770,08 %, у правому передсерді - 5,500,2, лівому - 4,960,76 %. Відносна площа кровоносних судин найменша у міжшлуночковій перегородці.
Отже, у добових теличок максимальна кількість кардіоміоцитів реєструється у міжшлуночковій перегородці і правому передсерді, а мінімальна - у лівому. Атипові кардіоміоцити у незначній кількості виявляються тільки в міокарді шлуночків і міжшлуночковій перегородці.
У робочих кардіоміоцитів превалює поздовжня посмугованість. Вставні диски виражені нечітко, ядра - овальної або округлої форми, що характерно для незавершених структур. Товщина кардіоміоцитів найбільша в міжшлуночковій перегородці, найменша - у лівому передсерді. У міокарді лівого шлуночка превалює діаметр робочих кардіоміоцитів і їхніх ядер, а об'єм - у міжшлуночковій перегородці. Найбільший відсоток пухкої волокнистої сполучної тканини спостерігається в міокарді правого передсердя, найменший - у лівому шлуночку і міжшлуночковій перегородці. Відносна площа кровоносних судин найбільша у лівому передсерді і шлуночку, а найменша - у міжшлуночковій перегородці.
У міокарді правого передсердя дослідних і контрольних 10-добових теличок кардіоміоцити так само, як і у добових, розташовуються окремими пучками, між якими знаходиться пухка волокниста сполучна тканина. Товщина робочих кардіоміоцитів у міокарді правого передсердя дослідних теличок на 3,34 % більша, ніж у контрольних (8,960,18 і 8,670,08 мкм), а довжина їх ядер практично однакова. Діаметр робочих кардіоміоцитів у теличок дослідної і контрольної груп змінюється аналогічно (Рис. 1). Поперечний діаметр ядер кардіоміоцитів у теличок дослідної групи збільшується на 6,45 %, порівняно з контрольними, а їх об'єм зростає лише на 1,12 %. У міокарді правого передсердя теличок дослідної групи спостерігається тенденція до збільшення площі поперечного перерізу ядер робочих кардіоміоцитів та їх цитоплазми. Ядра кардіоміоцитів як у дослідних, так і в контрольних теличок мають округлу або злегка видовжену форму. Ядерно-цитоплазматичне відношення має незначну різницю. Відносна площа пухкої волокнистої сполучної тканини більша у теличок контрольної групи, а кровоносних судин - у дослідних. Міокард лівого передсердя дослідних теличок утворений більш крупними поздовжньо і поперечно розташованими пучками кардіоміоцитов, що розділені тонкими прошарками сполучної тканини.
Провідні кардіоміоцити у міокарді передсердь теличок, як і у добових, не виявляються. Каріометричні структури міокарда лівого передсердя у теличок, вирощуваних із застосуванням дозованого примусового руху, мають тенденцію до збільшення. Ядра робочих кардіоміоцитів у теличок дослідної групи більш видовжені (коефіцієнт видовженості 2,310,05). Різниця в ядерно-цитоплазматичному відношенні преволює у теличок дослідної групи, що відображає інтенсивність розвитку і функціонального становлення клітин. До кінця неонатального періоду в дослідних теличок відносна площа пухкої волокнистої сполучної тканини зменшується на 12,26 % (P<0,05), а кровоносних судин збільшується на 7,72 % (P<0,01).
У міокарді правого шлуночка тільки у дослідних теличок робочі кардіоміоцити мають витягнуту форму, контактуючи меж собою "кінець у кінець", на що вказують ледь помітні вставні диски. У міокарді правого шлуночка дослідних теличок збільшується відносна площа кардіоміоцитів на 4,65, порівняно з правим, і на 8,85 % - з лівим передсердями, а в контрольних, відповідно, на 5,04 і на 8,91 %. У 10-добових дослідних і контрольних теличок виявляються провідні кардіоміоцити (11,080,25 і 11,270,14%). Товщина кардіоміоцитів у дослідних тварин (10,430,14 і 10,060,23 мкм), а довжина їх ядер (9,090,21 і 9,140,61мкм) не відрізняються, порівняно з контрольними. У міокарді передсердь і правого шлуночка дослідних теличок каріометричні показники збільшуються. Поперечний діаметр ядер робочих кардіоміоцитів більший на 4,69 %, площа поперечного перетину ядер - на 4,7, а цитоплазми на - 1,5 %, порівняно з контрольними теличками. Ядерно-цитоплазматичне відношення кардіоміоцітов шлуночків у дослідних теличок складає 0,620,03, проти 0,580,03 у контрольних. Відносна площа пухкої волокнистої сполучної тканини у теличок, вирощуваних із застосуванням дозованого примусового руху, зменшується на 6,4 %, а кровоносних судин зростає на 5,17 %, порівняно з тваринами, вирощуваних в умовах гіпокінезії. У робочих кардіоміоцитах лівого шлуночка дослідних теличок переважає поперечна посмугованість, а в контрольних - поздовжня. Відносна площа кардіоміоцитів у міокарді лівого шлуночка дослідних і контрольних теличок однакова, а провідних кардіоміоцитів на 0,66 % більша (P<0,05) у теличок контрольної групи (11,350,15 і 12,010,18%). Товщина робочих кардіоміоцитів у теличок дослідної групи збільшується на 10,71 % (8,720,41 і 7,460,71мкм), а довжина їх ядер - на 16,89 % (11,170,25 і 10,090,11 мкм, P<0,05).
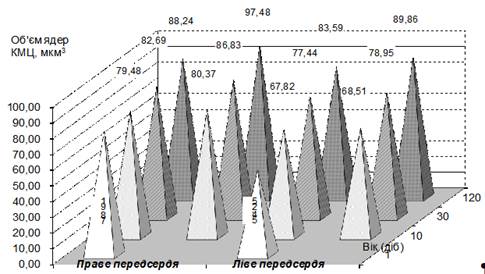
Рис.1. Динаміка об'єму ядер робочих кардіоміоцитів у передсердь теличок за різної рухової активністї
Поперечний діаметр ядер робочих кардіоміоцитів у дослідних теличок також на 6,96 % більший. У 10-добовому віці у теличок, вирощуваних із застосуванням дозованого примусового руху, зростає об'єм ядер кардіоміоцитів на 8,70 %, площа їх поперечного перерізу - на 7,0 %. Ядра кардіоміоцитів мають овальну форму. Коефіцієнт їхньої видовженості у дослідних теличок незначно більший, ніж у контрольних (1,960,05 і 1,890,11). Різниця в ядерно-цитоплазматичному відношенні в кардіоміоцитах дослідних теличок більша на 10,5 %, ніж у контрольних. Перші 10 діб життя теличок, вирощуваних із застосуванням дозованого примусового руху, зумовлюють більш інтенсивний ступінь диференціювання і функціонального становлення клітин та тканинних структур міокарда лівого шлуночка. До кінця неонатального періоду дослідних теличок у міокарді лівого шлуночка на 1,14 % зменшується відносна площа пухкої волокнистої сполучної тканини (8,260,42 і 7,120,28%, P<0,01) і кровоносних судин (2,830,1 і 3,110,12%, P<0,01). У міжшлуночковій перегородці теличок дослідної і контрольної груп виявляється найбільша відносна площа кардіоміоцитів. Товщина кардіоміоцитів у дослідних тварин однакова, порівняно з контрольними, але довжина їх ядер більша на 27,72 %. Нами не встановлено істотних розходжень каріометричних структур у міокарді міжшлуночкової перегородки. Проте у теличок дослідної групи спостерігається тенденція до збільшення поперечного діаметра кардіоміоцитів та їх ядер. Відносна площа пухкої волокнистої сполучної тканини у дослідних теличок зменшується (на 0,18%), а кровоносних судин збільшується на (0,39%). Максимальна кількість кардіоміоцитів реєструється в міжшлуночковій перегородці і правому шлуночку як дослідних, так і контрольних теличок. Провідних кардіоміоцитів у міокарді правого шлуночка і міжшлуночковій перегородці більше у контрольних теличок. У лівому шлуночку діаметр ядер робочих кардіоміоцитів, їх об'єм і площа поперечного перерізу більші у дослідних тварин. Найбільший відсоток пухкої волокнистої сполучної тканини в міокарді лівого передсердя у теличок, що вирощували в умовах гіподинамії, а найменший - у міжшлуночковій перегородці тварин, якім застосовували дозований примусовий рух. Відносна площа кровоносних судин превалює в лівому передсерді дослідних теличок, але вона значно менша в міжшлуночковій перегородці, порівняно з контрольними.
Застосування тваринам дозованого примусового руху зумовлює інтенсивне диференціювання клітин, що відображається в їх ядерно-цитоплазматичному відношенні. Міокард правого передсердя дослідних 30-добових теличок характеризується наростанням кількості кардіоміоцитів. У теличок дослідної і контрольної груп відносна площа кардіоміоцитів практично однакова, а кількість робочих превалює у дослідних. Товщина кардіоміоцитів правого передсердя дослідних теличок на 9,35 % більша, ніж у контрольних, а довжина їх ядер однакова. У теличок, вирощуваних із застосуванням дозованого примусового руху, переважають всі каріометричні показники кардіоміоцитів. Діаметр кардіоміоцитів у правому передсерді збільшується на 7,64, а діаметр ядер - на 20,88 %.
Наданий тваринам дозований примусовий рух сприяє підвищенню об'єму ядер на 4,76, а площі поперечного перерізу кардіоміоцитів - на 20,85 %. Ядра кардіоміоцитів у дослідних теличок мають видовжену форму, а в контрольних - овальну. За 30 діб життя у теличок дослідної групи відбувається зміна ядерно-цитоплазматичного відношення робочих кардіоміоцитів, що вказує на більш швидке диференціцювання клітин міокарда правого передсердя. У дослідних теличок на 0,67 % зменшується відносна площа пухкої волокнистої сполучної тканини, а кровоносних судин на 1,81% збільшується. Структура міокарда лівого передсердя дослідних теличок характеризується більш товстими пучками робочих кардіоміоцитів із чіткою поперечною посмугованістю.
У теличок місячного віку, вирощуваних із застосуванням дозованого примусового руху, превалює товщина кардіоміоцитів (на 5,32%), поперечний діаметр кардіоміоцитів (на 32,88%) і ядер (на 28,82%), порівняно з контрольними. Дозований примусовий рух сприяє збільшенню об'єму ядер кардіоміоцитів на 45,53%, площі поперечного перерізу ядер - 21,88, а цитоплазми - на 36,96 %. У дослідних теличок ядра стають більш видовженими (2,960,05 і 2,670,03; р<0,01). У міокарді лівого передсердя зменшується відносна площа пухкої волокнистої сполучної тканини, а кровоносних судин - зростає. У міокарді правого шлуночка дослідних теличок поперечна посмугованість стає чіткою, тоді як поздовжня виявляється незначно, відносна площа всіх кардіоміоцитів, як і робочих, дещо зменшується, а провідних стає на 1,03 % більше, ніж у контрольних. Товщина кардіоміоцитів і довжина їх ядер у дослідних тварин практично однакова. У правому шлуночці у теличок дослідної групи превалюють усі каріометричні показники робочих кардіоміоцитів (р<0,01). У дослідних тварин 30-добового віку відбувається збільшення об'єму ядер на 9,44 %, площі їх поперечного перерізу на 19,16, а цитоплазми - на 31,11 %, що приводить до зміни ядерно-цитоплазматичного відношення. У теличок, вирощуваних із застосуванням дозованого примусового руху, дещо зменшується відносна площа пухкої волокнистої сполучної тканини, а кровоносних судин, навпаки, зростає.
У міокарді лівого шлуночка серця у дослідних теличок кардіоміоцити утворюють суцільні поля. Відносна площа провідних кардіоміоцитів збільшується на 0,52 %. Застосування дозованого примусового руху протягом 30 діб теличкам сприяє збільшенню товщини кардіоміоцитів на 11,98, а довжини їх ядер - на 6,04 %. У дослідних тварин відбувається збільшення діаметра робочих кардіоміоцитів на 24,60 %, їх ядер - на 28,2 (P<0,05), а об'єму - на 9,3 %. Зростає площа поперечного перерізу ядер кардіоміоцитів на 28,19 і їх саркоплазми на 24,75 % (рис. 2).
Застосування дозованого примусового руху теличкам приводить до зменшення відносної площі пухкої волокнистої сполучної тканини і збільшення площі кровоносних судин. Робочі кардіоміоцити міжшлуночкової перегородки дослідних теличок мають чітку поперечну посмуговність, а в контрольних місцями ще зберігається повдовжня. У теличок дослідної групи зменшується відносна площа кардіоміоцитів, як і провідних, та зростає їх товщина (на 10,56%) і довжина їх ядер (на 11,18%). Дозований примусовий рух до 30-добового віку телят сприяє збільшенню діаметра робочих кардіоміоцитів на 30,23, їхніх ядер - на 24,64 %. У міжшлуночковій перегородці дослідних теличок превалює об'єм ядер кардіоміоцитів на 6,29 %, площа їх поперечного перерізу - на 24,5, а саркоплазми - на 30,27 %. Коефіцієнт видовженості ядра складає 2,360,07, а в контрольних теличок - 2,030,03 (р<0,01). У ядерно-цитоплазматичному відношенні робочих кардіоміоцитів істотної різниці не виявляється. У міжшлуночковій перегородці зустрічаються досить тонкі прошарки пухкої волокнистої сполучної тканини. У дослідних теличок на 0,75 % (р<0,05) збільшується відносна площа кровоносних судин. Отже, застосування дозованого примусового руху теличкам до 30-добового віку сприяє збільшенню товщини, довжини їх ядер і діаметра робочих кардіоміоцитів, особливо у міокарді лівого шлуночка і міжшлуночковій перегородці. Відносна площа кровоносних судин також превалює у тварин дослідної групи.
До 4-місячного віку зміни в апараті руху виявляються здебільшого у структурі тулуба, що пов'язано з розвитком внутрішніх органів. У міокарді правого передсердя дослідних і контрольних тварин поперечна посмугованість робочих кардіоміоцитів чітка. Ядра їх відносно великі, цитоплазма має щільне розміщення міофибрил. У дослідних теличок відносна площа всіх робочих кардіоміоцитів однакова, тоді як провідних - на 2,1 % більша.
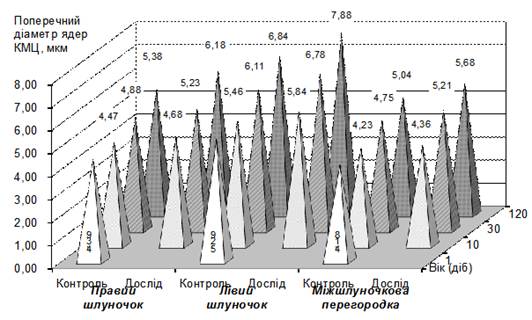
Рис.2. Динаміка поперечного діаметра ядер робочих кардіоміоцитів у шлуночках теличок за різної рухової активністі
У міокарді правого передсердя дослідних і контрольних теличок немає достовірної різниці між товщиною кардіоміоцитів і довжиною їх ядер. У 4-місячних теличок, вирощених із застосуванням дозованого примусового руху, відбувається збільшення каріометричних параметрів. Зростає поперечний діаметр кардіоміоцитів, особливо їх ядер (на 12,79%, р<0,01), та об'єм (на 10,47%, р<0,01, рис. 1, 2). Застосування тваринам дозованого примусового руху приводить до збільшення площі поперечного перерізу ядра на 12,78 і цитоплазми на - 5,11 %. У теличок дослідної групи відбувається зменшення відносної площі пухкої волокнистої сполучної тканини (на 2,33%) та збільшення кровоносних судин (на 2,21%). Міокард лівого передсердя дослідних і контрольних теличок характеризується компактно розміщеними робочими кардіоміоцитами, що утворюють своєрідні пучки, між якими виявляються різної товщини прошарки пухкої волокнистої сполучної тканини. У теличок дослідної групи товщина кардіоміоцитів зростає на 26,2, а довжина їх ядер - на 29,58 %. У кардіоміоцитах лівого передсердя відбувається збільшення всіх каріометричних показників. У дослідних теличок діаметр ядер кардіоміоцитів збільшується на 6,89, їх об'єм - на 7,62 %. Ядра кардіоміоцитів у дослідних тварин подовжені (2,840,05, 2,470,12, р<0,01). У теличок дослідної групи зменшується відносна площа пухкої волокнистої сполучної тканини (на 5,71%) і збільшується кровоносних судин (на 2,35%), порівняно з контрольними. У правому шлуночку теличок дослідної групи виявляється чітка поперечна посмугованість кардіоміоцитів та вставні диски між ними, а в контрольних ще превалює поздовжня. До кінця молочного періоду в теличок дослідної групи ще більше зростають товщина кардіоміоцитів (на 28,06%) і довжина їх ядер (на 13,78%). У правом шлуночку дослідних теличок поперечний діаметр кардіоміоцитів на 9,52, а їх ядер - на 14,86 % більший, ніж у контрольних. Найвищого значення досягає також об'єм ядер робочих кардіоміоцитів і площа поперечного перерізу цитоплазми (рис. 3). Відносна площа пухкої волокнистої сполучної тканини зменшується на 1,56 %, а кровоносних судин - на 2,19 % збільшується. Товщина робочих кардіоміоцитів зростає на 13,85 %, діаметр - 12,88 % а їх ядер - на 5,56 %, і 15,20 %.
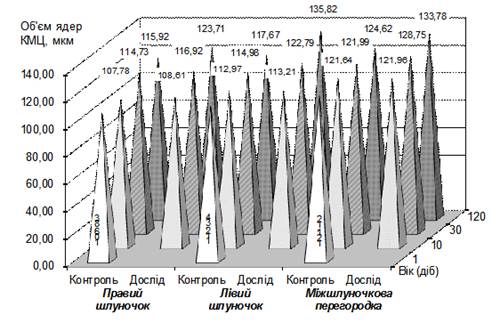
Рис.3. Динаміка об'єму ядер робочих кардіоміоцитів шлуночків теличок за різної рухової активності
Застосування дозованого примусового руху теличкам приводить до значного росту об'єму ядер кардіоміоцитів (на 15,42%). Площа поперечного перерізу ядра збільшується на 27,57 %, зростає різниця в ядерно-цитоплазматичному відношенні (1,350,005 і 0,940,005, р<0,001), порівняно з контрольними, що свідчить про більш інтенсивне диференціювання клітин міокарда. Відносна площа пухкої волокнистої сполучної тканини у дослідних теличок зменшується (на 2,74%), а кровоносних судин, навпаки, зростає (на 4,34%). Робочі кардіоміоцити міжшлуночкової перегородки дослідних теличок характеризується чіткою поперечною посмугованістю. За їх рахунок відбувається зменшення відносної площі всіх кардіоміоцитів. Товщина кардіоміоцитів у теличок дослідної групи зменшується на 4,6, а довжина їх ядер збільшується на 8,17 %. У міжшлуночковій перегородці діаметр кардіоміоцитів на 7,92 %, а їхніх ядер - на 12,69 % перевершує такі ж у контрольних. У дослідних тварин превалює об'єм ядер кардіоміоцитів на 7,35 %, площа поперечного його перерізу на - 11,75, а цитоплазми - на 7,88 %. На 0,87 % зменшується відносна площа пухкої волокнистої сполучної тканини, а кровоносних судин зростає на 1,83 %.
Корелятивні взаємозв'язки на органному рівні між товщиною стінки шлуночків, міжшлуночкової перегородки і поперечним діаметром кардіоміоцитів у теличок дослідної групи, як правило, тісні позитивні, а в контрольних - тісні зворотні або тісні позитивні.
Отже застосування дозованого примусового руху теличкам до кінця молочного періоду сприяє більш інтенсивному розвиткові робочих кардіоміоцитів, особливо міокарда шлуночків і міжшлуночкової перегородки. Зміна каріометричних параметрів робочих кардіоміоцитів у теличок дослідної групи супроводжується збільшенням поперечного їх діаметра та об'єму ядер у міокарді лівого шлуночка і міжшлуночковій перегородці, поперечна посмугованість стає чіткою. До 120-добового віку у них зменшується відносна площа пухкої волокнистої сполучної тканини, а кровоносних судин - збільшується.
Похожие статьи
-
У добових теличок масою тіла 34,62,86 кг абсолютна маса серця складає 247,519,8 г, а відносна - 0,720,05 %. Абсолютна маса кісткової системи досягає...
-
ВИСНОВКИ - Морфофункціональні особливості міокарда теличок за різної рухової активності
Морфофункціональні особливості серця телят залежать від віку, інтенсивності рухової активності, проявляючись на різних рівнях структурної організації...
-
Морфофункціональний стан, а також ріст і розвиток теличок, пов'язані з ростом і розвитком апарата руху. Разом з тим, складова його частина - кісткова...
-
Актуальність теми. Вирощування і утримання домашніх тварин спрямовано на максимальний розвиток їхньої продуктивності при мінімальних затратах. Проте...
-
Експериментальну частину досліджень проводили в умовах навчально-наукового виробничого тваринницького комплексу Кримського агротехнологічного...
-
Садівництво виникло дуже давно. Найдавніші описи садів і плодів трапляються в Ассірії та Вавілоні, Китаї, Індії, Греції. Давню історію має ця галузь і в...
-
Введение - Структурное состояние почв Пермского края и рекомендации по его улучшению
Способность почвы образовывать из механических элементов агрегаты носит название структурообразующей способности почвы, а совокупность получающихся в...
-
Общая характеристика предприятия Природно-климатические условия Географическое положение. ФГУП "Учхоз "Липовая гора" Пермской ГСХА имени академика Д. Н....
-
Изучая корневую систему трав, Саввинов (1936) установил, что оструктуривающее действие их сказывается более эффективно в почвенных зонах, наиболее...
-
Понятие о структуре почв - Структурное состояние почв Пермского края и рекомендации по его улучшению
Твердая фаза почвы состоит из механических элементов. Они смачиваются, взаимодействуют с жидкой фазой почвы, между собой и образуют агрегаты....
-
Состав и структура товарной продукции Структура выручки от реализации сельскохозяйственной продукции является одним из основных показателей...
-
Физические свойства почв - Структурное состояние почв Пермского края и рекомендации по его улучшению
Агрегатный состав пахотных почв и водопрочность агрегатов является важнейшим показателем плодородия. От него зависят такие агрофизические свойства почвы...
-
Морфологическая характеристика почв Морфологические признаки - это особый раздел почвоведения, характерезующий своим собственным предметом и методом...
-
Пермский район располагается на отложениях казанского яруса верхней Перми. Эти отложения состоят из красно-бурых (малиново-бурых) и коричнево-бурых...
-
Гидролитические условия - Структурное состояние почв Пермского края и рекомендации по его улучшению
В пределах города Перми протекают более 300 малых рек, речушек и ручьев. В левобережной части реки Кама, исследуемой территории г. Пермь, в микрорайоне...
-
Для забезпечення рентабельного виробництва свинини в умовах ринкової економіки весь технологічний процес повинен здійснюватися по чітко встановленій...
-
Рельеф - Структурное состояние почв Пермского края и рекомендации по его улучшению
Рельеф главный фактор перераспределения солнечной радиации и осадков. В зависимости от экспозиции и крутизны склона рельеф оказывает влияние на водный,...
-
Растительность - Структурное состояние почв Пермского края и рекомендации по его улучшению
Изучаемая часть землепользования ФГУП УОХ "Липовая гора" относится ко 2-му району южно-таежных елово-пихтово лесов подзоны южной темно-хвойной тайги...
-
Климат Территория города Пермь (микрорайон "Липовая гора") находится в четвертом агроклиматическом районе, подрайоне б. Данный район по...
-
Заключение - Структурное состояние почв Пермского края и рекомендации по его улучшению
1) Основными коренными породами на территории г. Пермь и его окрестностях являются отложения Шешминского горизонта Уфимского яруса Пермской системы:...
-
У комплексі негативних антропогенних чинників особливе місце за масштабами й небезпекою впливу посідає аеротехногенне забруднення. Техногенне надходження...
-
По данным работы [12] среднестатистические потери зерна при скашивании составляют в среднем 40-50% и иногда достигают 80% от общих потерь за жаткой,...
-
Особливості стану лісових насаджень Фастівського району Державне підприємство "Фастівське лісове господарство" розташоване на площі 18596 гектарів на...
-
Фактори, які впливають на пророщування зерна - Технологічні особливості процесу пророщування солоду
Основними факторами, які впливають на пророщування являються температура, вологість і доступ кисню (аерація). Найбільший вплив надає температура....
-
Після замочування зерно передають разом із водою в солодовню, де і відбувається процес пророщення. Мета пророщування - а) накопичення і переведення в...
-
Бонитировка почв - Структурное состояние почв Пермского края и рекомендации по его улучшению
Работы по бонитировке почв в России были начаты давно, еще в XV в. Научные Основы оценки почв (бонитировки) разработаны в конце прошлого столетия В. В....
-
Сорти яблуні за термінами споживання плодів у свіжому вигляді діляться на літні, осінні і зимові [1]. У літніх сортів знімна стиглість плодів збігається...
-
1. Яблуня - одна з найдавніше окультурених плодових рослин зони помірного клімату. Культура яблуні має велике значення у світовому плодівництві,...
-
Ми вже говорили про те, що догляд за подібним садом значно простіший, ніж за багатими насадженнями сильнорослих плодових дерев, але варто відзначити і...
-
Ботанічна характеристика: Соя належить до роду, який, за даними F. J.Herman (1962), поділяється на три підроди (submenus) Leptocyamus, Glycine і Soja. За...
-
Одна з головних проблем у технології No-till це бур'яни. Соя має досить довгий гербокритичний період - 45-50 днів. Триває він від появи першого...
-
Недоліки карликових дерев - Особливості вирощування карликових сортів яблунь на території України
На жаль, є й деяка кількість недоліків при вирощуванні карликового саду. Серйозні початкові витрати. Звісно ж, при посадці карликових садів потрібно...
-
Ранній вступ у плодоносіння. Це дуже важливий показник, як для промислового, так і для приватного садівництва, адже плодові дерева на ділянці ми...
-
Яблуня належить до ботанічного роду Malus з порядку розоцвіті (Rosales), родини розанні ( Rosaceae Suss ), підродини яблуневі (Romideae Focke). Вона...
-
Агропроизводственная группировка почв представляет собой объединение в более крупные группы видов и разновидностей почв, близких по агрономическим...
-
Вступ - Особливості вирощування карликових сортів яблунь на території України
Культура яблуні в Україні - важлива галузь сільського господарства, яка дає населенню цінні продукти харчування - плоди. Свіжі яблука в нашій державі...
-
Агрохимические свойства характеризуются показателями: емкость катионного обмена, сумма поглощенных оснований, обменная и гидролитическая кислотность,...
-
Вид - Колорадський жук - Leptinotarsa decemlineata Say Родина листоїди - Chrysomelidal Рід твердокрилі - Coleoptera Поширений скрізь. До середини 60х рр....
-
Ботанічна характеристика та біологічні особливості кукурудзи Ботанічна характеристика кукурудзи У світовому рільництві, у тому числі й в Україні,...
-
Висновок - Оцінка лісових насаджень Фастівського лісгоспу
Лісистість України становить 15.9%. Україна займає 8 місце в Європі за площею лісів та 6 місце за запасами деревини. Загальна площа лісів-- понад 10 млн...
Структурно-функціональні особливості міокарда теличок за різної рухової активності - Морфофункціональні особливості міокарда теличок за різної рухової активності