Биосинтез и деградация эндоканнабиноидов - Антигипоксическое и нейропротекторное действие N-арахидоноилдофамина при моделировании острой гипоксии in vivo и in vintro
Каннабиноиды синтезируются и реализуются локально On demand (по требованию). Таким образом, уровень их генерации не является постоянным, а кинетика их деградации является основным фактором их активности. В естественных условиях, как полагают, синтез анандамида происходит путем ферментативного гидролиза фосфолипазой D мембранного предшественника - N-арахидоноил фосфотидилэтаноламина (Schmid et al., 1983; Deutsch, Chin, 1993). N-арахидоноил фосфотидилэтаноламин формируется путем ферментативного переноса арахидоновой кислоты из sn-1 положения мембранного фосфотидилхолина на аминогруппу фосфотидилэтаноламина (Cadas et al., 1997; Sugiura et al., 2002). Никаких конкретных трансацилаз, катализирующих данный процесс, на сегодняшний день не идентифицировано, однако некоторым исследователям удалось клонировать специфичную для N-арахидоноил фосфотидилэтаноламина фосфолипазу D (Okamoto et al., 2004). Возможны и другие варианты синтеза анандамида. Так, например, в желудке секреторная фосфолипаза А2 может катализировать конверсию N-ацил фосфотидилэтаноламина в N-ацил лизофосфотидилэтаноламид. Далее под действием лизофосфолипазы D образуются соответствующие N-ацилэтаноламиды, в том числе, анандамид (Sun et al., 2005). Другой альтернативный путь был описан в RAW246.7- макрофагах, где происходит гидролиз N-арахидоноил фосфотидилэтаноламина до фосфоанандамида с участием фосфолипазы С и дальнейшим действием фосфатазы (Liu et al., 2006).
Для N-ADA предполагались два возможных пути биосинтеза. Первый возможный путь - это прямое сопряжение арахидоновой кислоты и дофамина, а другой - через метаболизм предполагаемого N - арахидоноилтирозина (NA-тирозин) (рис. 3).
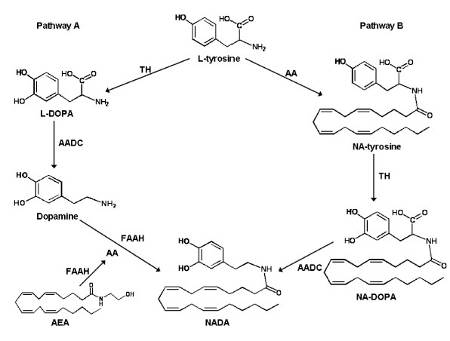
Рисунок 3 Два предполагаемых метаболических пути биосинтеза N-ADA (Hu et al., 2009)
Показано, что NA-тирозин не является промежуточным продуктом в ходе биосинтеза N-ADA, который синтезируется в первую путем ферментативного сопряжения арахидоновой кислоты с дофамином (Hu et al., 2009). Хотя это сопряжение, вероятно, включает в себя комплекс ферментов, показано прямое участие гидролазы амидов жирных кислот в биосинтезе N - ADA.
Эндоканнабиноиды не хранятся в синаптических везикулах, как другие нейротрансмиттеры. Напротив, они, благодаря своей липидной природе, через постсинаптическую мембрану попадают в синаптическую щель, где связываются с соответствующими рецепторами, часто расположенными пресинаптически.
Таким образом, ЭК могут выступать в качестве сигнальных молекул. Дальнейшие сигнальные пути, опосредуемые CBR, приводят к ингибированию реализации нейромедиаторов, главным образом ГАМК или глутамата, таким образом, ретроградно модулируя синаптическую передачу (Gуmez-Ruiz et al., 2007).
Чтобы прекратить влияние ЭКС, эндогенные лиганды СВR, как и другие нейротрансмиттеры, должны быть инактивированны. Эта инактивация происходит в 2 этапа: ЭК должны быть транспортированы в клетку, возможно, специальной транспортной системой, а затем гидролизованы под действием специальных ферментов (Bari et al., 2006). Анандамид гидролизуется до двух компонентов (арахидоновой кислоты и этаноламина) под действием гидролазы амидов жирных кислот (ГАЖК, FAAH), а 2-арахидоноилглицерол - до арахидоновой кислоты и глицерина под действием моноацилглицероллипазы (МАГЛ, MAGL) (Dinh et al., 2002).
В 1994 г. было обнаружено, что захват анандамида обратно в клетку происходит путем облегченной диффузии (Di Marzo et al., 1994), и что этот процесс зависит от ряда факторов (температуры, чувствительности к субстрату, насыщаемости). Также была показана его независимость от ионных градиентов и гидролиза АТФ, однако возможно влияние оксида азота (Maccarrone et al., 2000). Хотя в последние годы было предпринято немало попыток идентификации специфического транспортера ЭК, его природа до сих пор остается не известной, что порождает все новые споры о его существовании (Glaser et al., 2003). На сегодняшний день, как правило, принято считать, что движение анандамида через клеточную мембрану - процесс насыщаемый, а также предложены различные механизмы поглощения анандамида, в том числе облегченная диффузия (Fegley et al., 2004), простая диффузия с ГАЖК-опосредованным расщеплением анандамида, простая диффузия с внутриклеточной секвестрацией анандамида, или эндоцитоз (McFarland et al., 2004). В отличие от анандамида, об обратном захвате 2-AG практически ничего не известно. Некоторые авторы предполагают, что он попадает в клетку опосредованно специальным транспортером, другие - что использует транспортер анандамида (Bisogno et al., 2000), либо это происходит путем простой диффузии (Beltramo, Piomelli, 2000).
В дополнение к основным ферментам, в процессе утилизации анандамида и 2-AG могут участвовать различные оксигеназы, например, липоксигеназа, цитохром Р450, циклооксигеназа, превращая ЭК в соответствующие соединения простагландина (van der Stelt et al., 2002). Физиологическая роль данных реакций пока не ясна.
Похожие статьи
-
N-арахидоноилдофамин Все агонисты CBR принято подразделять на растительные (фитоканнабиноиды), эндогенные и синтетические. Среди фитоканнабиноидов (рис....
-
РОФ - функционально разнородная группа белков, способных быстро изменять свою концентрацию при различных нарушениях гомеостаза независимо от природы и...
-
Дарвин о видообразовании. Микроэволюция - Сущность эволюционной теории Чарльза Дарвина
Возникновение новых видов Дарвин представлял как длительный процесс накопления полезных изменений, увеличивающихся из поколения в поколение. Мелкие...
-
Спектры фотобилогического действия - Фотохимические превращения ДНК. Люминесцентные метки и зонды
Изучение характера действия и силы действия светового излучения, относящегося к определенному световому диапазону, обычно начинают с определения спектра...
-
Химический состав микробов. Вода составляет основную массу микробной клетки - в капсульных бактериях ее больше, в бациллах меньше. В Aerobacter aceti...
-
Ген и его строение - Биосинтез белков. Ген и его роль в синтезе белков
Ген представляет собой элементарную единицу функции наследственного материала. Это означает, что фрагмент молекулы ДНК, соответствующий отдельному гену и...
-
Фагоциты как центральное звено неспецифической защиты организма На современном этапе развития медицинской науки иммунофагоцитарная система...
-
Введение - Биосинтез белков. Ген и его роль в синтезе белков
Способность клеток поддерживать высокую упорядоченность своей организации в хаотичной Вселенной зависит от генетической информации, которая реализуется,...
-
Этиология и патогенез острого холецистита: - Острый холецистит
Воспалительный процесс в стенке желчного пузыря может быть обусловлен не только микроорганизмом, но и определенным составом пищи, аллергологическими и...
-
В каждом организме (животном, растительном или микробной клетке) все время происходят сложные процессы превращения веществ. Велика роль в этих процессах...
-
Действие на организм и токсичность - Метаболизм этилового спирта в организме
Этиловый спирт может поступать в организм несколькими путями: при приеме внутрь, при внутривенном введении, а также через легкие в виде паров с вдыхаемым...
-
Поражаемая система, Механизм действия - Токсикология мышьяка
Различают следующие типы отравлений мышьяком, которые могут явиться причиной летальных исходов: Паралитический, возникающий при огромных дозах мышьяка;...
-
Этиология Возбудитель заболевания относится к дерматотропным поксвиру-сам. Поражает различные отделы кожи, в том числе лица и век. Передается...
-
Диагностика, Инструментальное обследование пациентов при ИМ - Острый инфаркт миокарда
Лабораторные методы диагностики инфаркта миокарда: Џ Анализ крови Џ Биохимический анализ крови Џ Кровь на миоглобин Резорбционно-некротический синдром -...
-
Этиология и патогенез - Лечебные и профилактические мероприятия при гипоксии внутриутробного плода
Причинами острой гипоксии являются: 1. Неадекватная перфузия крови к плоду из материнской части плаценты (например при нарушении дыхания роженицы) . 2....
-
Взаимопревращения пластид - Пластиды: строение, происхождение, взаимопревращения
В эволюционном смысле первичным, исходным типом пластид являются хлоропласты, из которых при расчленении тела растений на органы произошли пластиды...
-
Аденозинтрифосфорная кислота (АТФ) состоит из органического основания аденина (I), углевода рибозы (II) и трех остатков фосфорной кислоты (III)....
-
Таким образом, управление жизнедеятельностью организма путем организации ФС гомеостатического и поведенческого уровней обладает рядом свойств,...
-
Фотохимические реакции - Фотохимические превращения ДНК. Люминесцентные метки и зонды
ФОТОХИМИЧЕСКИЕ РЕАКЦИИ - химические превращения, протекающие под действием света в видимой и ультрафиолетовой области спектра. Еще в античные временя...
-
Гемодиализ - Заболевание почек и печени
Гемодиализ не может полностью заменить собой почки, но с помощью этого метода больные могут нормально жить и работать, несмотря на почечную...
-
Экспрессия генов - Экспрессия генов
Экспрессия генов -- это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный...
-
Возбудитель чумы - Частная микробиология
Специфическая профилактика не разработана. Для лечения используют антибиотики широкого спектра действия. Таксономия. Y. enterocolitica относится к отделу...
-
Механизмы апоптоза - Радиационный апоптоз и некроз клеток облученного организма
Развитие и функционирование многоклеточных организмов подразумевает равновесие между синтезом и распадом молекул, их окислением и восстановлением,...
-
Метод микрофигурии - Методы изучения клетки
С помощью микроманипулятора отдельные части клетки можно удалять, добавлять или каким-то образом видоизменять. Крупную клетку амебы удается разделить на...
-
1) [+]2, 3, 6, 9, 10, 12 2) [-]1, 3, 4, 5, 6, 11 3) [-]2, 5, 7, 8, 10, 12 4) [-]1, 4, 5, 7, 8, 11 Как называются птицы, птенцы которых вылупляются из...
-
ПАТОГЕНЕЗ - Острый панкреатит, отечная форма
Основной теорией патогенеза острого панкреатита является ферментативная. Основным доказательством этой теории является экспериментальное получение...
-
СОСАЛЬЩИК ЛЕГОЧНЫЙ - Paragonimus westermani, P. ringeri - возбудители парагонимоза - антропозооноза. Географическое распространение - районы...
-
Крапивница и отек Квинке, Побочные действия местных анестетиков - Лекарственная аллергия
А. В патогенезе крапивницы и отека Квинке, вызванных Аспирином , могут участвовать разные механизмы. Б. Аспирин и другие НПВС вызывают обострение и...
-
Реализация функциональной активности фагоцитирующих клеток находится, в том числе, под контролем различных отделов нервной системы и регуляторных...
-
Причины - Анемия при беременности
Основной причиной развития ЖДА являются кровопотери различной природы. Они нарушают существующее в организме равновесие между поступлением и выведением...
-
Диагностика - Анемия при беременности
Оценка тяжести заболевания основывается на показателях содержания гемоглобина, уровня гематокрита, концентрации железа в плазме крови (в норме 13 - 32...
-
Основным запасным источником энергии в организме являются жиры. По мере необходимости жиры из жирной ткани поступают в Виде неэстерифицированных...
-
Аутоиммунные процессы, Ожирение и питание - Изменение при сахарном диабете
На возможность значения аутоиммунного процесса (Nerup J. et all, 1978) в развитии диабета указывают ряд данных: 1) присутствие в островках поджелудочной...
-
Первая помощь при нарушении дыхания - Физиология и гигиена органов дыхания у детей
Инородные тела в дыхательных путях . Опасная ситуация, связанная с закупоркой дыхательных путей, может возникнуть даже в самой будничной обстановке -...
-
ЯЗВЕННАЯ БОЛЕЗНЬ - Организация сестринского процесса при осложнениях язвенной болезни
Язвенная болезнь - хронически рецидивное заболевание, склонное к прогрессированию с вовлечением в патологический процесс других органов системы...
-
В эпидемиологии развития инфекции имеют значение: 1. возраст 2. географическое положение 3. социально - экономический статус 4. профессия Инфицирование...
-
ДИАГНОСТИКА - Организация сестринского процесса при осложнениях язвенной болезни
1. рентгенологические исследования 2. эндоскопические исследования 3. УЗИ 4. ФГДС ЛЕЧЕНИЕ Цель медикаментозной терапии - облегчение боли, заживление...
-
Заболевание распространено на всех континентах. Болеют в основном собаки, кошки, пушные звери, кролики, морские свинки, хомячки и птица. Регистрируются у...
-
Антибиотики - вещества, избирательно угнетающие жизнедеятельность микроорганизмов, т. е. АБ активны только против микробов при сохранении...
-
- трофобласт и эмбриобласт - первичная полоска и Гензеновский узелок + эпибласт и гипобласт - эктодерма и энтодерма - синцитиотрофобласт и цитотрофобласт...
Биосинтез и деградация эндоканнабиноидов - Антигипоксическое и нейропротекторное действие N-арахидоноилдофамина при моделировании острой гипоксии in vivo и in vintro