Етапи еволюції роду Stipa і формування степів
Етапи еволюції роду Stipa і формування степів
Степовий біом (разом із преріями і пампасами) на земній кулі займає близько 6 % площі суходолу і є одним із головних у функціонуванні біосфери. Степи акумулюють енергетичні ресурси в найбагатших за вмістом гумусу чорноземних грунтах, що забезпечують високі врожаї сільськогосподарських культур. Разом з тим вони нещадно експлуатуються, а тому деградація степового біому та скорочення площі природних екосистем нині настільки значні, що про збереження їхньої цілісності та всього різноманіття вже не йдеться. Ступінь розораності територій степів в Україні сягає 87--96 %, значні їхні площі деградовані, а таких, що мають природну структуру, -- всього 1--3 %. Загальна втрата запасів гумусу варіює у межах 4,3--3,5 %, що призводить до виснаження їх енергетичного потенціалу (Medvedev, 2002).
Степи були колискою високого видового біотичного різноманіття, в результаті чого сформувалися специфічні ксерофітні біотопи трав'янистого типу (злаковники) та пасовищної фауни, що історично тісно взаємопов'язані. З метою збереження вцілілих клаптиків і виконання екосистемами властивих їм функцій, з-поміж іншого, важливим є дослідження історії формування степових таксонів та екосистем. Ці питання перебували в полі зору багатьох відомих учених, викликали гострі дискусії, і хоча вони далекі від остаточного вирішення, але сприяють розвитку теорії степознавства й еволюції природи. Використання нових підходів, сучасних методів досліджень відкриває невідомі сторінки історії формування степів, що наближає нас до уявлення цілісної картини цього складного і багатогранного процесу. Водночас знання такої історії важливе для розробки заходів зі збереження степів.
З цього погляду перспективним є дослідження триби Stipeae Dumort. як найтиповішого елементу степової біоти. Метою роботи було встановлення філогенетичних відносин між представниками триби Stipeae, оцінка таксономічної структури триби на основі молекулярних даних (ДНК) і мікроморфологічних ознак, виявлення біогеографічних закономірностей та часу формування представників триби, напрямків їх розселення; визначення етапності розвитку степів у зв'язку з кліматичними змінами й еволюцією інших природних складових, зокрема зоокомпонентів.
Об'єкти та методи досліджень
Для повної репрезентації філогенетичних груп використано гербарний матеріал Інституту ботаніки імені М. Г. Холодного НАН України (KW), Ботанічного інституту імені В. Л. Комарова РАН, Росія (LE) та Смітсонівського інституту, США (USNC). Ампліфікацію ДНК видів триби Stipeae проведено в лабораторії Смітсонівського інституту й Інституту ботаніки в Барселоні (Іспанія). Для аналізу були обрані десять маркерів із хлоропластного геному -- rpl32-trnL, rps16- trnK, rps16 intron, trnK-matK, matK, trnh-psbA, trnL-F, ccsA, ndhF і ndhA; інтергенний регіон із нуклеарного геному -- ITS і два гени низької копії At103 і AroB з нуклеарного геному. Протоколи й умови ампліфікації, а також розроблені ДНК - праймери описані в роботі К. Ю. Ромащенка та ін. (Romaschenko et al., 2011, 2014). Усього проаналізовано 396 зразків світових Stipeae, з них 111 належать до групи справжніх ковилів, серед яких більшість українських видів. Загалом отримали 1961 генетичний сиквенс, які розміщені в базі Генбанку NCBI та Базі даних ДНК-баркодингу.
Клонування та пряма ампліфікація At103 гена здійснені в Лабораторії Інституту ботаніки Барселони, а клонування та пряма ампліфікація гена AroB -- у Смітсонівському інституті за тією самою методикою. Для побудови філогенетичного дерева застосовано парсімоніальний аналіз (програма PAUP 4b.10), аналіз максимальної подібності (програма GARLI 2.0) та аналіз Баєсівської інференції (програма MrBayes 3.1.2). Для статистичної підтримки клад філогенетичного дерева використовували бутстреп аналіз і параметри постеріорної вірогідності. Для дослідження давньої гібридизації між видами слугував генетичний ретикулярний аналіз (програма Splitstree 4.11.3). Період диверсифікації головних філогенетичних груп розраховано методом молекулярного годинника із аналізом Баєсівської інференції (програма BEAST 1.5.1), що відображає різницю темпів мутацій між генетичними маркерами та філогенетичними групами. З метою побудови генеалогічної схеми за набором гаплоїдів AroB використано програму TCS 1.21, що працює за принципами статистичної парсимонії.
Результати досліджень та їх обговорення Філогенія та генеалогія роду Stipa. Методика молекулярного годинника дала змогу визначити періоди виникнення головних груп у Stipeae
Та швидкість генетичних змін у межах різних еволюційних ліній (рис. 1). Приблизні дати утворення головних філогенетичних груп Stipeae такі. Євразійська група виникла 21 млн років тому. В її межах рід Stipa з'явився на початку міоценового кліматичного оптимуму та близько 11 млн років тому розділився на дві групи, які переважно охоплюють азійські та європейські Stipa. Як і очікувалося, дати виникнення Ptilagrostis Griseb., за аналізом послідовності нуклеарної та хлоропластної ДНК, не збігаються (3 і 6 млн років відповідно). Це свідчить про самостійне походження хлоропластних ліній, які старші за нуклеарні. Зіставлення результатів аналізу молекулярного годинника для нуклеарних і хлоропластних даних допомогло реконструювати генетичну еволюцію базових груп триби Stipeae у міоцені--пліоцені. Ми припускаємо існування на початку міоцену на території Азії трьох головних нуклеотипів -- це ксеро-ірано - туранський (proto-Ampelodesmos, Psammochloa та Neotrinia), мезо-ірано-туранський (proto-Trikeraia тa proto-Oryzopsis) і східноазійський (proto - Patis I). Відразу після кліматичного оптимуму, в середньому міоцені, геноми Orysopsis Michx. і Trikeraia Bor. відмежувалися. Близько 13 млн років тому відбулася інтрогресія геному Oryzopsis до proto-Patis І і формування геному proto-Patis II. Приблизно 13 млн років тому Oryzopsis мігрував до Америки через Берингію, і цей таксон повністю зник з території Євразії. Близько 9 млн років тому відбулося відділення Patis obtusa (Stapf ex Oliv.) Romasch., P. M. Peterson &; Soreng і формування геному para-Patis I. Приблизно 5--6 млн років тому могла статися зворотна міграція proto-Ptilagrostis з Американського континенту в Азію, в результаті чого відбулась інтрогресія американського нуклеотипу до Para-patis I тa рroto-Trikeraia, що призвело до формування геному рara-Рatis II і сучасної Trikeraia. Через патернальне зворотне схрещування з батьківськими формами утворилися дві групи видів Ptilagrostis: одна має гаплотип Trikeraia (більшість існуючих видів Ptilagrostis), а друга -- гаплотип Patis (Ptilagrostis alpina (F. Shmidt) Sipliv. і P. porteri (Rydb.) WA. Weber). Останній 1,7--2,9 млн років тому, в час осциляції льодовиків, мігрував до Америки. Дещо раніше (4,3 млн років тому) розділилися Patis coreana (Honda) Ohwi тa P. racemosa (Sm.) Romasch., P. M. Peterson &; Soreng, що теж мігрував до Америки. Отже, генетичний обмін відбувався через ірано-туранські території та поширювався в західному напрямку. Ксеротичні ірано-туранські таксони не брали участі в цьому геномному обміні. Таким чином, отримані дані свідчать, що до Берингової протоки було дві хвилі азійсько-американської міграції Stipeae -- 13 тa близько 5 млн років тому (риа 1). При цьому група Pulcherrimae монофілетична, тоді як Pennatae -- парафілетична (рис. 1).
Stipa степ палеозоокомплекс
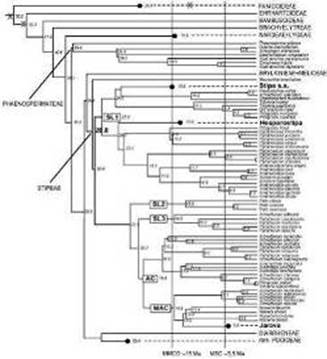
Рис.1
Попри те, що групи азійських і європейських Stipa розділилися раніш як 10 млн років тому, генетична різноманітність у межах цих груп є дуже низькою. Це свідчить про швидку генетичну радіацію сучасних Stipa. Серед низки тестованих хлоропластних і нуклеарних маркерів єдиним, що відобразив певну диференціацію філогенетичних груп, був маркер AroB. Встановлено, що всі види азійських Stipa мають дві копії гена, одна з яких належить до Ptilagrostis. Цим пояснюється наявність перехідних морфологічних ознак у видів даної групи, які часто відносили до Ptilagrostis, тому ми запропонували розглядати їх у ранзі підроду Pseudoptilagrostis Tzvelev, до якого належать S. capillacea Keng, S. regeliana Hack., S. subsesilliflora (Rubr.) Roshev., S. purpurea Griseb., S. penicillata Hand.-Mazz., S. rohmooiana Noltie тa S. aliena Keng. Решта видів об'єднана в subgen. Stipa, де базовими є секції Barbatae Junge, Smirnovia Tzvelev, Leiostipa Dumort. та група азійських видів роду Stipa, що охоплює S. brevifolia Griseb. i S. bungeana Trin. Давньосередземноморська секція Barbatae виявилася базовою для решти видів. Вона поділяється на дві клади -- середземноморську, що містить S. barbata Desf. і S. lagascae, та cередньоазійську, яка охоплює S. meyeriana (Trin. et Rupr.) Grossh., S. hohenackeriana Trin. et Rupr. і S. badachshanica Roshev. Усі інші види Stipa, котрих іноді відносили до цієї секції, належать до Leiostipa або формують самостійні лінії (рис. 1). На наступному етапі відгалузилась азійська група (види S. bungeana тa S. brevifolia), що, як і види subgen. Pseudoptilagrostis Tzvelev, отримали додаткові копії гена AroB. Це свідчить про генетичний обмін із родом Ptilagrostis. Дві монофілетичні групи Smirnovia тa Leiostipa є сестринськими, вони об'єднані в спільну кладу. Типовий вид групи Leiostipa -- S. capillata L. Він має специфічну структуру генеалогічного сиквенсу, яка стабільна в усіх зразках -- від Західного Середземномор'я до України, Кавказу та Казахстану. До Leiostipa також належать S. macroglossa P. A. Smirn., S. krylovii Roshev., S. baicalensis Roshev., S. daghestanica Grossh., S. grandis P. A. Smirn., S. karatabinica і S. areptana A. Beck. До групи Smirnovia відносять S. desertorum (Roshev.) Ikonn., S. caucasica Schmalh., S. longiplumosa Roshev., S. drobovii (Tzvelev) Chern. і S. iskanderkulica (Tzvelev) Chern. До однієї клади зі Smirnovia та Leiostipa належать види, які формують незалежні лінії, -- S. spiridonovii Roshev., S. richteriana
Kar. &; Kir., S. himalaica Roshev., S. brauneri (Pacz.) Klokov тa S. lessingiana Trin &; Rupr.
Отже, останні два види, поширені в Україні, належать не до секції Barbatae, а до клади Smirnovia -- Leiostipa.
У свою чергу, в складі секції Barbatae виокремлюється група Arabica (Barbatae II) (S. arabica Trin. &; Rupr., S. trichoides P. A. Smirn. i S. turkestanica Hack.), яка заслуговує на виділення в окрему секцію (Romaschenko et al., 2014). Саме вона є сестринською щодо українських (європейських) секцій Pennatae тa Pulcherrimae.
Нажаль, маркер AroB не надає достатньої інформації для підтвердження або заперечення існування групи Pennatae як філогенетичної одиниці, для цього потрібен пошук інших маркерів. До групи Pennatae відносять поширені в Україні S. tirsa Stev., S. lithophila P. A. Smirn. (а не до Pulcherrimae, як вважалося раніше), S. disjuncta Klokov, S. graniticola Klokov, S. borysthenica Klokov ex Prokudin, S. asperella Klokov &; Ossycznjuk, S. pontica P. A. Smirn. і S. maeotica Klokov &; Ossycznjuk. Останні три види формують окрему кладу, що суперечить виділенню групи Atlanticae, до якої вносили S. asperella P. A. Smirn., S. iberica Martinovsky тa S. atlantica P. A. Smirn. з опушеною адаксіальною поверхнею листкової пластинки (Klokov, 1975; Martinovsky, 1966). З іншого боку, S. asperella, S. pontica тa S. maeotica мають жорстке опушення на абаксіальному боці листкової пластинки, що є кращою філогенетичною ознакою. Незважаючи на велику морфологічну подібність S. disjuncta, S. graniticola, S. pennata (S. joannis Celak.) і S. borysthenica, остання вирізняється специфічною структурою генетичного сиквенсу, яка витримується для всіх зразків із материкової частини України, Криму, Кавказу, тому сумніву щодо її видового статусу не має бути. Відзначимо, що з екологічного погляду цей псамофітний вид чітко відрізняється від інших степових.
Молекулярний аналіз показав, що генетична структура групи Pulcherrimae в Україні не є гомогенною. Кримські види відокремлені та мають сильніші зв'язки з кавказькими, ніж з рештою східно-, центрально - та західноукраїнських видів. До кримської групи належать S. heterophylla Klokov, S. oreades Klokov, S. glabrinoda Klokov, а з Кавказу -- S. araxensis Grossh. i S. pulcherrima. Паннонська S. transcarpatica Klokov виявляє тісні генетичні зв'язки з піренейською S. iberica Martinovsky. Окремі клади формують балканські та південноєвропейські S. eriocaulis Borbas, з України -- степові з S. pulcherrima, що генетично відмежовані від кавказьких S. dasyphylla і S. zalesskii Wilensky. Отже, остання належить не до групи Pennatae, а до Pulcherrimae.
Встановлено, що Stipa adoxa Klokov &; Ossycznjuk i S. syreistschikovii PA. Smirn. формують окремі лінії, а Stipa oreades має гібридне походження.
Для з'ясування генеалогічних зв'язків в окремих групах Stipa здійснено аналіз статистичної парсимонії. Метод дає можливість проаналізувати послідовність накопичення мутацій; види, які розміщуються вище, вважаються молодшими, ніж ті, що нижче. За цим аналізом доведено, що групи Pennatae i Pulcherrimae є дериватними. Базовий у межах Pulcherrimae -- гаплотип S. syreistschikovii, а всі інші -- похідні. Незрозумілим залишається гаплотип S. eriocaulis. Наймолодші за генеалогічним аналізом європейські види з Іспанії (S. iberica) тa Карпат (S. transсarpatica). У групі Pennatae базовою для S. maeotica i S. pontica виступає S. asperella (рис. 1).
Встановлено, що, на відміну від груп Pennatae та Рulcherrima, всі інші групи Stipa характеризуються великою кількістю нереалізованих гаплотипів. Це свідчить про високий ступінь зникнення таксонів у ході еволюції та значну тривалість еволюційних процесів. Порівняно з цими групами в групах Pennatae і Pulcherrimae нереалізованих гаплотипів майже немає, і генеалогічна схема загалом репрезентує швидку радіацію видів, для оцінки яких потрібен пошук інших маркерів. Це означає, що, незважаючи на широке розповсюдження, Pennatae тa Pulcherrimae є молодими групами і швидко еволюціонують у зв'язку з розширенням зони аридних територій, кліматичними змінами плейстоцен--голоцену, формуванням зоокомплексів пасовищного типу, кочівництвом і скотарством.
ВплИв еколого-кліматичних змін і зоокомплексів на формування степів. Підходи до технології їх збереження. Використання молекулярного годинника на основі молекулярно-генетичного аналізу дає змогу встановити часові (реперні) точки відліку появи тих чи тих таксонів. Однак багато аспектів, пов'язаних із формуванням новітніх близькоспоріднених таксонів, усе ще залишаються невідомими. У цьому плані поряд із пошуком відповідних молекулярно-генетичних маркерів перспективним є застосування класичних методів флорогенетичного аналізу (Vulf, 1944; Popov, 1963; Kamelin, 1979) у комплексі з сучасними даними палеонтології, палеокліматології, палеоекології (Velychko, 1973; Vereshhagin, 1979; Lavrenko, 1981). Це дає можливість відобразити характер кореляції між зміною зовнішніх чинників та еволюцією ознак як способів адаптації до навколишнього середовища і визначити основні моменти формування степів, зокрема, простежити зв'язок із еволюцією палеозоокомплексів (рис. 2).
На початку палеогену (палеоцен--нижній еоцен), коли з'явилися злаки, сформувався диноцератовий палеозоокомплекс, який утворювали древні ссавці, представлені великими тваринами. Вони населяли ліси та заболочені місцевості і розвивалися в умовах субтропічного та тропічного клімату всієї Північної півкулі. Вважається, що в цей період зональність означеної території недостатньо виражена, бо диференціація фауни не проявляється. Панували великі травоїдні носорогоподібні (диноцерати, амінодонти, гіракодонти), предки слонів (пантоламбди), тапірів (гірахуси, ореодонти), копитних (кондиляртри), зокрема коней (гіракотерії, орогіппуси), тощо (Bazhanov, Erofeev, 1971). В еоцені пра-Євразія відділилася від пра-Північної Америки, а в середньому еоцені Тургайське (Сибірське) море розділило Євразію на західну та східну частини, що спричинило територіальну ізоляцію континентів і подальшу еволюцію ссавців та рослин. У верхньому еоцені--нижньому олігоцені диноцератовий палеозоокомплекс змінився на бронтотерієвий (бронтотерії, амінодонти, ентелодонти, халікотерії, прістотерії, предки коней -- палеотерії), що населяли вологі ландшафти тургайського типу теперішньої Євразії (Bazhanov, Erofeev, 1971). У цей період вичленилися древні представники триби Pooidae, які, очевидно, були лісовими елементами тургайської флори.
У середньому та верхньому олігоцені в ландшафтах усе ще переважали лісові та болотні типи, що заселялися великими жуйними ссавцями індрикотерієвого палеозоокомплексу (індрикотерії, гіракодонти, амінодонти, тапіроїди, халікотерії, ентелодонти, антракотерії тощо). У складі злаків виділяється триба Stipeae та інші.
З переходом від палеогену до неогену (верхній олігоцен--міоцен) у Північній півкулі вже спостерігається диференціація клімату за ознаками термічності та континентальності, що спричинилося підняттям суші (Гімалаїв) і висиханням величезного Тургайського моря на території сучасної Західносибірської низовини і, як наслідок, -- сформувалися пралісовий і прасаванний типи біоти. Деревний ярус прасаван мав вигляд ксеротичних рідколісь, утворених видами східноазійського походження (Diels, 1910), що сприяло інтенсивному розвитку трав'яного покриву, зокрема злаковників. Диференціація злаків відбувається на рівні сучасних родів у трибі
Stipeae --Achnatherum, Piptatherum і в інших трибах, анцестральні форми яких були все ще лісовими видами -- представники Poa L. (sect. Homalopoa Dumort.), Festuca L. (subgen. Drymanthele V. I. Krecz. &; Bobrov, Schedonorus (P. Beauv.) Peterm.), Dacty - lis L.; однак формуються таксони і ксерофітного, геліофітного типів палеосаван. Еволюція ссавців відбувається в напрямку скорочення жуйних і розширення розмаїття тварин анхітерієвого палеозоокомплексу, пристосованих до швидкого бігу, які живилися сухими жорсткими травами.
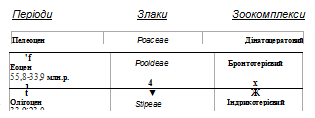
Рис. 2. Схема коеволюції степових злаків і палеозоокомплексів Fig. 2. Scheme of coevolution of steppe grasses and paleozoocomplexes
Цей анхітерієвий міоценовий палеозоокомплекс, який, окрім видів саван, охоплював і лісових предків коней (Parahippus-Meryhippus-Dinohippus), носорогів(Rhinoceratoidea),мастодонтів
(Gomphoterium), оленів-мунтжаків (Muntiacus muntjak), свиноподібних, газелей, халікотеріїв, динотеріїв, гризунів тощо, значною мірою впливав і на еволюцію рослинного світу. Взаємозв'язок між зоо - та флористичною складовими визначав характер і напрямок коеволюції (рис. 2). Саме з початком неогену біоморфи трав'яного типу починають виступати як домінанти і виникають анцестральні форми сучасних типів екосистем. Щодо тодішніх палеосаван, то в майбутньому на їхній основі у тропічній зоні формувалися савани, а в помірній -- степи та прерії.
У цей період, 10--20 млн років тому, в трибі Stipeae виникли основні групи рослин, які мали довгі та короткі клітини епідерми, та окремі роди, зокрема рід Stipa (15 млн років тому).
У міоцен--пліоцені (2--12 млн років тому) тривали подальша інтенсивна еволюція тварин відкритих просторів саванового типу, зокрема копитних, і формування гіпаріонової фауни, що мігрувала на великі території, очевидно, через сезонні зміни клімату та припинення вегетації рослин у посушливі періоди. У складі гіпаріонової фауни відзначені конеподібні (Plesiohippus, Hipparion), носороги (Aceratherium, Elasmotherium), хоботні (Anancus, Mastodon), антилопи (Gasellopsira), верблюди (Paracamelus), олені (Eucladoceros, Cervus, Arvernoceros, Libralces), жирафи (Palaeotragus) (Korotkevich 1988; Iordanskij, 2001), значне розмаїття регіональних зоокомплексів (кучурганський, хапровський, молдавський, таманський, тираспольський), що відображають різні стації живлення (Logvynenko, 2002; Titov, 2008). Рештки викопної фауни та характер пилкових спектрів із Artemisia L. і Chenopodiaceae опосередковано свідчать про ксеричний характер природних умов зі збереженням великих лісових масивів у долинах рік, що сприяло еволюції та розселенню представників біоти. Зокрема, досягли свого розквіту порожньорогі (Bovinae), які були предками овець і кіз та зіграли велику роль у розселенні й еволюції ковилів. У цей період у межах триби Stipeae відбуваються родова диференціація, їхня міграція, зокрема в Північну Америку та в зворотний бік. 5--7 млн років тому скорочувалися морські басейни Сарматського, а пізніше (2--3 млн років тому) -- і Меотичного морів. Це спричинило посилення континенталізації та потужну експансію аридного біотичного комплексу зі Східної до Центральної Азії в Європу, але до Атлантики. Цей ксеротичний ірано-туранський комплекс, в основі якого були групи Barbatae, Smirnovia, Leostipa, цілковито ізолювався.
Як вважали Є. М. Лавренко та В. С. Говорухін (Lavrenko, Govorukhin, 2000), наприкінці пліоцену відбулася заміна саванового комплексу степовим, хоча елементи степового типу існували і раніше у високогір'ях; власне степова зона як географічне, зональне явище вичленувалася значно пізніше, в голоцені.
Перехід від пліоцену до плейстоцену (2--4 млн років тому) характеризувався наростанням похолодань, що зумовило зміну гіпаріонової фауни та появу тварин з довгою шерстю. У цей період з'явилися коні (Equus), великорогі олені (Megaloceros giganteus), еласмотерії (Elasmotherium sp.), мамонти (Mammutus trogontherii, M. primigenius та ін.), верблюди (Camelidae sp.), шерстисті носороги (Coelodonta antiquitatis), степові бізони (Bison priscus), тури (Bos primigenius), вівцебики (Ovibos moschatus), сайги (Saiga tatarica s. l.). Дані молекулярної філогенії засвідчують, що предки сучасних коней, які з'явилися 4--4,5 млн років тому, вівцебиків (Ovibos) та овець (рід Ovis, родини Bovidae)-- азійського походження (Campos et al., 2010; Orlando et al., 2013). Останні були представлені в цьому фауністичному комплексі 3 млн років тому і через Північну Азію, Беринговий міст мігрували, з одного боку, до Америки, а з другого -- до Європи, де їхнє розселення було пов'язане з гірськими масивами (муфлони, архари) (Shnirelman, 1980; Rezaei et al., 2010). Очевидно, це сприяло подальшій диференціації і в складі триби Stipeae.
Важливим етапом в еволюції ксеротичного, аридного біотичного комплексубув період плейстоцену (2 млн -- 12 тис. років тому), коли спостерігався відступ льодовиків. Так, на думку В. Я. Сергіна та С. Я. Сергіна (Sergin, Sergin, 1993), наростанняльодовиківспричинило похолодання та посилення зволоження, а їхній відступ супроводжувався ксеризацією за холодного клімату, яка потім змінюваласяпотеплінням.
Наступний етаппотеплінняпов'язаний
Із зростанням гумідизації, зволоженням, похолоданням і знову -- наступом льодовиків. Такі зміни суттєво впливали на еволюцію степової біоти. Очевидно, її формування відбувалося від палеосаванового типу через прастеповий, тундро - степовий до степового, коли з похолоданням наростала і континенталізація, а теплолюбні деревні види саван змінювалися холодостійкими пребореальними тундро-степовими. Для таких біотопів характерний спорово-пилковий комплекс із переважанням пилку Pirns L. sp. (90 %), а з трав'янистих видів -- Chenopodiaceae (30 %), Artemisia (20 %), Rosaceae і Fabaceae (до 10 %), хоча не вони визначали характер рослинності, а були індикаторами наростання ксеротичної складової. У ній важливу роль відігравали злаки та складноцвіті, пилок яких зберігається гірше. У фауністичному аспекті для плейстоцену характерний мамонтовий палеозоокомплекс, що сформувався на основі гіпаріонової фауни і був адаптований до континентальних умов, бо тварини мали густу та довгу шерсть (Zazula et al., 2003; Levchenko, 2004; Burroughs, 2005; Smirnova, 2006). Суттєвий вплив на формування степів і тундростепів пов'язують з в'юрмським періодом, що тривав понад 50 тисячоліть, під час якого тундрові та степові комплекси поєднувалися. Такі тварини, як сайга, проникали далеко на північ, а вівцебики -- на південь, до зони сучасного лісостепу (Vangengeim, 1977). Н. К. Верещагін (Vereshhagin, 1979) наводить для цього комплексу близько 40--50 видів копитних, гризунів та інших тварин, які безпосередньо харчувалися травами, що, звісно, впливало на формування ценозів. Анатомо-морфологічні ознаки видів роду Stipa свідчать, що для утворення дернин потрібні витоптування та випасання, а перенесення плодів відбувається вітром і в довгій шерсті тварин. Його представниками були шерстистий мамонт (Mammutus primigenium), шерстистий носоріг (Coelodonta antiquitalis), степовий бізон (Bison priscus), тур (Bos primigenius), вівцебик (Ovibos moschatus), у харчуванні багатьох із яких переважали злаки й осоки (Ukraintseva, 2002; Iordanskij, 2010). Наприкінці плейстоцену з'явилися великі барани (Ovis cf. ammon). Саме такі кліматичні умови та фауністичний комплекс визначали переважання екосистем пасовищного типу ("мамонтові степи"), сприятливого для лучних і степових злаків, зокрема ковили (рис. 4). При цьому формувалися два комплекси: північний холодний, власне мамонтовий (Mammutus, Equus caballus, Bison priscus, Coelodonta, Saiga tatarica) і південний, з наявністю (Bison, Equus hemionus, Coelodonta, Ovis cf. ammon). Якщо з першим пов'язано утворення тундро-степів, аналогом яких Є. М. Лавренко (Lavrenko, 1981) вважає степові угруповання Якутії, де в ценозах домінують Festuca (F. lenensis Drob.,), Koeleria Pers. (K. cristata (L.) Pers., Carex L. (C. pediformis C. A. Mey., C. duriuscula C. A. Mey.), Agropyron cristatum (L.) P. Beauv., Helictotrichon krylovii (Pavl.) Henrard, Stipa (група Leiostipa), то з останнім -- прастепів, де основу становлять елементи південніших територій Монголії та Середньої Азії.
На таких відкритих просторах росли й дерева у вигляді рідколісь (Pinus L., Larix Mill., Betula L.), хоча їхній пилок і становить дуже великий відсоток у спорово-пилкових спектрах (Grichuk, Grichuk, 1960), однак на основі сучасних методів інтерпретації таких спектрів доведено, що панували тоді трав'янисті, а не лісові ландшафти (Konovalov, Ivanov, 2007). Цілком логічно, що межі між ними не були різкими, траплялися рідколісся від ксерофітного до мезофітного типів (Didukh, 2007). Саме з останніми ми пов'язуємо існування анцестральних елементів майбутніх степових і лучних угруповань, що мають генетичні зв'язки, простежені нами на прикладі родів Poa, Festuca, Dactylis, Carex, із прабореальними, а не неморальними комплексами (Didukh, 1992).
Плейстоцен характеризувався чергуванням чотирьох льодовикових і міжльодовикових епох, що спричинило відповідні зміни природних умов. Діагностувати специфіку кожної з них щодо еволюції степової флори сьогодні ще неможливо. Молекулярно-генетичний аналіз відображає лише деякі моменти. В льодовикові епохи на прильодовикових територіях відклалися потужні товщі лесу, які слугували ареною для заселення ксерофітних видів. Тут відбувались інтрогресивна гібридизація та радіальна еволюція рослин. Саме останнє утруднює застосування молекулярних маркерів. Тому в цьому випадку використовують опосередковані дані, наприклад, особливості екологічних умов, географічні зв'язки між таксонами, характер диз'юнкції їхніх ареалів тощо. Так, встановлені на основі молекулярних маркерів тісні зв'язки між кавказькими та кримськими видами Stipa, з одного боку, і карпатськими й іберійськими -- з другого, паралельно з диз'юнкціями інших таксонів свідчать про найпізніший зв'язок між кримсько-кавказьким і європейським ексклавами у рисс-в'юрмі, а між ними -- в міндель-риссі. Отже, рисський льодовик спричинив розрив між Південно-Західною Європою та Евксинським регіоном. У свою чергу, найпізніший зв'язок між Кримом і Кавказом міг бути лише в рисс-в'юрмі. Це спостерігається у структурі ареалів багатьох видів різних систематичних груп (роди Thymus, Stipa, Eremurus M. Bieb., Helianthemum Gray., Jurinea Cass.) і в їхніх відмінностях на расовому рівні, яким деякі автори надавали рангу видів (Klokov, 1973). Ці зв'язки переривалися не лише через кліматичні, а й інші суттєві зміни комплексу факторів, наприклад, потужних відкладів лесу, піскових морен, що визначали розвиток екосистем й окремих складових біоти і грунтотворення. На підтвердження диз'юнкцій цього періоду С. Фірбас (Firbas, 1949) наводить факти зростання кількох видів роду Helianthemum у Скандинавії, які вважає в'юрмськими реліктами.
Саме з пострисським, а пізніше -- з поств'юрмським періодами пов'язане поступове формування псамофітного степового комплексу, характерними компонентами якого є Centaurea L. ser. Arenariae (Hayek) Dostal (C. borysthenica Gruner, C. arenaria M. Bieb. ex Willd.), ser. Margaritaceae Klokov (C. margaritacea, C. margaritaalba Klokov, C. protomargaritacea Klokov, C. paczoskii Kotov &; Klokov, C. konke Klokov, C. appendicata Klokov, C. donetzica Klokov), Thymus ser. Thymiciliani (Th. borysthenicus Klokov &; Des.- Shost., Th. odoratissimus Mill.), Agropyron Gaertn. (A. dasyanthum Ledeb., A. tanaiticum Nevski, cristatum subsp. sabulosum Lavrenko, A. cimmericum Nevski), Dianthus L., Otites Adans., Jurinea. (Lavrenko, 1936, Klokov, 1981). Очевидно, до цього комплексу слід віднести й Stipa borysthenica. Цей псамофітний комплекс пов'язаний не тільки з боровими терасами, що сформувалися в часи пізнього плейстоцену (поств'юрму), ай із більш ранніми пострисськими піщаними відкладами, які на правих, підвищених, берегах рік Донецької та Середньоруської височин залягають на крейдяних відкладах і не перекриваються лесовими.
Другою такою ареною інтенсивного видоутворення були крейдяні та гранітні відслонення в степовій зоні. Саме тут описано багато дрібних видів і рас, що мають генетичні зв'язки зі псамофітними та степовими видами (Stipa graniticola Klokov, Otites hellmannii (Claus) Klokov, O. dolichocarpa Klokov, O. maeotica Klokov, Onosma graniticola Klokov, Thymus kajmijussicus Klokov &; Des.-Shost.). Водночас у цих умовах знайшли прихисток елементи так званої гісопової флори, що генезисно пов'язана з Передньоазійсько - середньоазійським флористичним центром.
Наступний етап інтенсивного розвитку степових екосистем і їхньої флори припадає на голоценовий період. Саме в голоцені могла сформуватися степова зона як географічне явище, характерною ознакою якої є домінування дерновинних злаків (Lavrenko, 1940; Lavrenko, Govorukhin, 2000). Значною мірою це пов'язано з розвитком кочівництва та скотарства. Так, на основі палеозоологічних даних встановлено, що одними з перших тварин, приручених у часи палеоліту-- неоліту (8--12 тис. років тому), були вівці, предком яких виступав дикий гірський баран -- муфлон, що живе в умовах суворого аридного клімату (Shnidelman, 1980; Rezaei et al., 2010). Після винищення фауни пасовищного типу інтенсивний розвиток кочівництва, скотарства, зокрема вівчарства, спричинив формування степової рослинності, в угрупованнях якої домінувала Stipa (Avdeev, 1948). Доказом цього є те, що ані коротка шерсть корови чи коня, а лише вовна вівці чи кози може забезпечити перенесення зернівки, яка угвинчується у вовну і відламується. Цьому сприяє і гвинтоподібно заточений носик зернівки, коліновигнута вість, що нагадує буравчик.
Як показують експериментальні дані, для збереження та відновлення степових ковилових угруповань у Національному парку "Віденський ліс" організовано випас овець, які інтенсивно виїдають рослини до основи дернини, ущільнюють грунт, а після випасу, стресової для ковили ситуації, наступного року вона інтенсивно квітне. Отже, одним із факторів втрати степової компоненти в наших степах є відсутність випасу, зокрема овець.
Намагання зберегти степову основу за відсутності випасання призводить до того, що надмірні енергетичні запаси не відчужуються, відтак екосистема зазнає відповідних змін, які відомі з класичних законів ендоекогенетичних сукцесій. Рослинні угруповання не відтворюють собі подібних, а характеризуються певним зміщенням, напрямок і швидкість якого регулюється впливом зовнішніх чинників (Didukh, 2008). Навіть за досягнення певної рівноваги щодо зовнішніх умов і стійкого стану вони продовжують змінюватися; в результаті їхня першооснова руйнується і переходить в якісно інший стан, який неможливо передбачити через складну взаємодію і кумулятивний ефект впливу зовнішніх факторів, що продовжують трансформуватися. Ксерофітні степові дерновинні угруповання змінюються короткокореневищними злаками, потім -- різнотрав'ям, нітрофільними видами, з'являються чагарники, дерева, формуються лігнозні типи угруповань (Tkachenko, 2004) .
Щоб зберегти існуючий стан екосистеми, необхідно відчужувати певні запаси енергії, не давати змоги їй накопичуватися (Didukh, 2014). Раніше функцію відчуження виконували дикі тварини, які до освоєння людиною населяли степи у великій кількості, а згодом тривав потужний випас свійських тварин (Avdeev, 1948). Але в степових заповідниках цей випас був припинений, відтак почалися суттєві зміни екотопів.
Особливої гостроти ця проблема набула для степових заповідників ще в 60-ті роки ХХ ст., коли в умовах абсолютного заповідання вони почали втрачати степову компоненту, з'явилися лучні види, кущі, дерева і т. д. Відомі вчені -- Г. І. Білик,
В. В. Осичнюк і В. С. Ткаченко -- запропонували замінити випас сінокосінням, яке би стримувало швидкість змін, але це кардинально не поліпшило ситуації. Сьогодні залучнення та закущення степових ділянок у заповідниках триває швидкими темпами. Це спричинено і кліматичними змінами: адже в степовій зоні за останні сто років кількість річних опадів збільшилася на 100 мм. Тому в заповідниках ми втрачаємо степові екосистеми, паралельно там скорочуються і популяції рідкісних видів.
Отже, ми вважаємо, що в розробці технології щодо режимів заповідання, збереження екосистем слід опиратися на глибоке знання історичного формування біоти у взаємозв'язку зі зміною навколишнього середовища. Використання молекулярно-генетичного аналізу в комплексі з анатомо-морфологічними даними, еколого - флорогенетичними підходами дає важливі результати і є основою для розробки менеджмент - плану організації степових заповідників (Didukh, 2014).
Висновки
Отримані молекулярно-генетичні дані допомогли встановити періоди виникнення головних груп у Stipeae та швидкість генетичних змін еволюційних ліній. Приблизна дата появи головних філогенетичних груп триби Stipeae -- 16--21 млн років тому. Рід Stipa з'явився на початку міоценового кліматичного оптимуму і близько 11 млн років тому розділився на дві групи, які переважно охоплюють азійські та європейські Stipa, а 5 млн років тому, наприкінці міоцену, виникла "справжня" Stipa.
На основі молекулярно-генетичного аналізу доведено, що генетична структура групи Pulcherrimae в Україні не є гомогенною. Кримські види перебувають ізольовано і мають сильніші зв'язки з кавказькими, ніж з рештою українських видів. З'ясовано, що S. zalesski належить не до групи Pennatae, а до Pulcherrimae. Stipa adoxa та S. syreistschikovii формують окремі лінії, а Stipa oreades -- гібридного походження. S. lessingiana і S. brauneri належать до групи Smirnovia--Leiostipa.
На основі статистичної парсимонії встановлено, що групи Pennatae i Pulcherrimae -- похідні від Barbatae. Базовим у межах Pulcherrimae є гаплотип S. syreistschikovii. Наймолодшими видами за генеалогічним аналізом визнано види з Іспанії (S. iberica) тa карпатські S. transсarpatica.
У групі Pennatae базовою для S. meotica i S. pontica є S. asperella, а S. borysthenica розглядається як самостійний вид. Молекулярно-генетичний аналіз у комплексі з флорогенезисними, палеонтологічними, палеокліматичними, палео - екологічними даними відображають характер кореляції між зміною зовнішніх умов та еволюцією ознак як способів адаптації до навколишнього середовища. Це дало змогу визначити основні моменти формування степів від палеосаван (у неогені), тундро-степів (у плейстоцені) до степів у голоцені. Такий розвиток саваново - степових елементів пов'язаний із коеволюцією зоокомплексів пасовищного типу.
Формування сучасних степів зумовлено розвитком кочівництва та скотарства, а для збереження ковилових ценозів необхідний помірний випас овець.
Список літератури
Stipa степ палеозоокомплекс
- 1. Avdeev V. D. Vozniknoveniestepey vZakame, Kazan: Tatgo - sizdat, 1948, 51 pp. [Авдеев В. Д. Возникновение степей в Закамье. - Казань: Татгосиздат, 1948. -- 51 с.]. 2. Bazhanov V. S., Erofeev V. S. Smena teriofaun Kaynozoya basseyna verkhnego Irtysha na fone tektonicheskogo rezhima i klimaticheskoy obstanovki. In: Yskopaemaya fauna y flora Tsentralnoho y vostochnoho Kazakhstana, Alma-Ata: Nauka, 1971, vol. 5, pp. 63--66. [ Бажанов В. С., Ерофеев В. С. Смена териофаун Кайнозоя бассейна верхнего Иртыша на фоне тектонического режима и климатической обстановки // Ископаемая фауна и флора Центрального и Восточного Казахстана/ Отв. ред. Б. С. Кожамкулова. -- Алма-Ата: Наука, 1971. - Т 5.- С. 63-66]. 3. Bilyk G. I. Luchni stepy. In: Roslynnist URSR: Stepy, kam'yanysti vidslonennya, pisky, Kyiv: Naukova Dum - ka, 1973, pp. 33-94. [Білик Г. І. Лучні степи // Рослинність УРСР: Степи, кам'янисті відслонення, піски / Відп. ред. А. І. Барбарич. - К.: Наук. думка, 1973. - С. 33-94]. 4. Campos P. F., Willerslev E., Sher A., Orlando L., Axelsson E., Tikhonov A., Aaris-Sorensen K., Greenwood A. D., Kahlke R.-D., Kosintsev P, Krakhmalnaya T, Kuznetsova T, Lemey Ph., MacPhee R., Norris C. A., Shepherd K., Suchard M. A., Zazula G. D., Shapiro B., Gilbert T. P. Ancient DNA analyses exclude humans as the driving force behind late Pleistocene musk ox (Ovibos moschatus) population dynamics, Proceedings of the National Academy of Sciences, 2010, 107(12): 5675-5680. 5. Cialdella, A. M., Sede S. M., Romaschenko K., Peterson PM., Soreng R. J., Zuloaga F. O., Morrone O. Phylogeny of Nassella (Stipeae, Pooideae, Poaceae) Based on Analyses of Chloroplast and Nuclear Ribosomal DNA and Morphology, Systematic Botany, 2014, 39 (3): 814 -828. 6. Didukh Ya. P Rastitelnyi pokrov Gornogo Kryma (struktura, dinamika, evolyutsiya i okhrana), Kyiv: Naukova Dumka, 1992, 256 pp. [Дидух Я. П. Растительный покров Горного Крыма (структура, динамика, эволюция и охрана). - Киев: Наук. думка, 1992. - 256 с.]. 7. Didukh Ya. P. Rol ekologii ta geografii roslin u rozrobtsi problem rozvitku roslinnogo pokrivu. Etyudi fitoekologii, Kyiv: Aristej, 2008, pp. 27-64. [Дідух Я. П. Роль екології та географії рослин у розробці проблем розвитку рослинного покриву. Етюди фітоекології. - К.: Аріс - тей, 2008. - C. 27- 64]. 8. Didukh Ya. P. Stepnoy bull., 2014, 40: 8-10. [Дидух Я. П. Что мы должны охранять в степных заповедниках? // Степной бюлл. - 2014. - 40. - С. 8-10]. 9. Didukh Ya. P. Chto takoe lesostep? In: Botanika imikolohiya: Sovremennye horizonty, Kyiv: Akademperiodika, 2007, pp. 40-53. [Дидух Я. П. Что такое лесостепь? // Ботаника и микология: Современные горизонты: Сб. тр. / Отв. ред. А. А. Созинов. -- Киев: Академпериодика, 2007. - С. 40-53]. 10. Diels L. Genetische Elemente in der Flora der Alpen, Botanische Jahrbucher fur Systematik, Pflanzengeschichte undPflanzengeographie, Beiblatt, 1910, 44(4): 7-46. 11. Firbas S. Spat - und nacheiszeitliche Waldgeschichte Mitteleuropas nordlich der Alpen. In: Allgemeine Waldgeschichte, Jena, 1949, Bd. 1, 480 pp. 12. Grichuk M. P., Grichuk V. P. O prilednikovoy rastitelnosti na territorii SSSR. In: Periglyatsialnye yavleniya na ter - ritorii SSSR, Moscow: Izd-vo MGU, 1960, pp. 66-100. [Гричук М. П., Гричук В. П. О приледниковой растительности на территории СССР // Перигляциальные явления на территории СССР. - М.: Изд-во МГУ, 1960 - C. 66-100]. 13. Iordanskij N. N. Evolyutsiya zhizni, Moscow: Akademia, 2001, 425 pp. [Иорданский Н. Н. Эволюция жизни. - М.: Академия, 2001. - 425 с.]. 14. Kamelin R. V. Kukhystanskyi okruh hornoy Sredney Azyi, Leningrad: Nauka, 1979, 117 pp. [Камелин Р. В. Ку - хистанский округ горной Средней Азии. - Л.: Наука, 1979. - 117 с.]. 15. Klokov M. B. Psammofilnye floristicheskie kompleksy na territorii USSR (opyt analiza psammofitona). In: Novosti sistematiki vysshikh i nizshikh rasteniy, Kyiv: Naukova Dumka, 1981, pp. 90-150. [Клоков М. В. Псаммофильные флористические комплексы на территории УССР (опыт анализа псаммофитона) // Новости систематики высших и низших растений. - Киев: Наук. думка, 1981. - С. 90-150]. 16. Klokov M. V., Osychnyuk V. V Kovyli Ukrainy. In: Novosti systematiki vysshykh i nizshikh rasteniy, 1975. Kyiv: Naukova Dumka, 1975, pp. 7-92. [Клоков М. В., Осычнюк В. В. Ковыли Украины // Новости систематики высших и низших растений. - Киев: Наук. думка, 1976. - С. 7-92]. 17. Konovalov A. A., Ivanov C. N. Izyestiya Chelyab. nauch. tsen - tra. Ser. Biologiya, 2007, 1(35): 125-130. [Коновалов А. А., Иванов С. Н. Новый способ реконструкции палеоклимата по спорово-пыльцевым спектрам // Изв. Челяб. науч. центра. Сер. Биол. - 2007.- Вып. 1(35). - С. 125-130].
Похожие статьи
-
Палеомалакологічні, які і інші палеонтологічні та палеогеографічні дослідження, проводяться у декілька етапів. Першим, зазвичай, є підготовчий етап,...
-
У залежності від умов проживання усі молюски можна поділити на морські і континентальні. Серед останніх виділяють власне континентальні (наземні) та...
-
Визначення черепашок молюсків проводиться шляхом порівняння знайдених екземплярів з ілюстраціями і описами в відповідних визначниках і атласах. На цій...
-
Волинська лесова височина -- класичний район розповсюдження лесів. Леси утворюють тут майже суцільний покрив(за винятком річкових заплав і крутих схилів...
-
Волинська височина, за П. Цисем [31], є північною геоморфологічною підобластю Волино-Подільської височини. Її продовженням у Польщі є Люблінська...
-
З наведених вище переліку фауни молюсків кожного із розрізів та результатів її аналізу можна зроби наступні висновки про природні умови часу формування...
-
Просторово-часові особливості формування системи розселення Харківської області
У статті виявлено просторово-часові особливості формування системи розселення Харківської області, проведено кластерний аналіз за демографічними...
-
При проведенні польових робіт важливе значення має організація першої (невідкладної) медичної допомоги. Перша медична допомога представляє собою...
-
Вимоги інструкції є обов'язковими для всіх структурних підрозділів університету, організаторів експедицій, для всіх співробітників, студентів,...
-
Рештки викопних молюсків вивчалися у різних відкладах здавна. Але тільки з другої половини ХІХ століття такі дослідження стали приймати науковий...
-
При дослідженні розрізу нами було відібрано 10 проб для палеомалакологічного аналізу: одну з горизонту А горохівського викопного грунтового комплексу,...
-
Загалом походження лесу Волинської височини більшість вчених пов'язують з плейстоценовими зледеніннями. У результаті кліматичних змін, викликаних...
-
Зупинимось більш детально на умовах існування прісноводних молюсків, оскільки їх також часто знаходимо у четвертинних відкладах, як у тих, що мають водне...
-
Для проведення магістерського дослідження застосовувалося ряд методів, серед яких слід виділити малакологічний. Він використовувався для отримання,...
-
Постановка проблеми. На сьогоднішній день існуючі системи розселення мають безліч недоліків, серед яких незбалансований соціальний, економічний,...
-
У всіх випадках проведення польових робіт заборонено: * розводити вогнища на відстані ближче 10 м від наметів і на відстані 100м від складу...
-
Вплив снігового покриву на клімат - Вплив підстильної поверхні на формування клімату
Сніговий покрив утворюється лише у певних кліматичних умовах, тобто він є продуктом клімату. Але в той й же час сніговий покрив впливає на клімат. При...
-
Четвертинний покрив Волинської височини сягає потужності 30м. Він побудований, в основному, відкладами лесово-грунтової серії і тільки на заході височини...
-
У даному розрізі нами опробувались лише леси підгоризонту 2г, де було виявлено: Pupilla sp. - 2 екземпляри; Sphaerium sp. - 2 екземпляри; Pisidium milium...
-
Вплив грунту та рельєфу на кліматоутворення - Вплив підстильної поверхні на формування клімату
На суходолі тепло вглиб передається внаслідок молекулярної теплопровідності. У товщі води тепло передається значно ефективнішим методом, а саме завдяки...
-
Фауна викопних молюсків середньо-верхньоплейстоценових лесів на території Волинської височини вивчалась у розрізах, які подано у таблиці 3.6, а...
-
Розріз Торчин - опорний розташований у кар'єрі цегельного заводу селища Торчин Волинської області. Геоморфологічно - це північна частина розчленованої...
-
Розріз Коршів - опорний знаходиться за 18 км південно-західніше м. Луцька, біля с. Ново-Коршів, в великому кар'єрі по видобутку сировини для цегельних...
-
Вступ, І, Клімат. Загальна інформація - Вплив підстильної поверхні на формування клімату
Тема роботи: у цій роботі я підніматиму тему кліматотвірних чинників та впливу підстильної поверхні на клімат зокрема. Мета роботи : поглибити знання про...
-
Висновок, Список використаних джерел - Вплив підстильної поверхні на формування клімату
Отже, підстильна поверхня і дійсно є одним із визначальних чинників кліматоутворення. Ми узагальнили знання про кліматотвірні чинники та процеси,...
-
Вплив рослинного покриву на клімат - Вплив підстильної поверхні на формування клімату
Через те, що Земля має форму геоїда та отримує різну кількість сонячної радіації на різних широтах, через нерівність рельєфу на нашій планеті...
-
Волинська височина розташована у межах південної частини Волинської і Рівненської, північної частини Львівської областей, а також (частково) Люблінського...
-
Кліматотворчі процеси -- атмосферні процеси, що визначають характер клімату в межах тієї чи іншої області чи всієї Землі (теплообіг, вологообіг, загальна...
-
Вплив водного простору на кліматоутворення - Вплив підстильної поверхні на формування клімату
Взагалі вода засвоює сонячного тепла більше, ніж суходіл, оскільки альбедо води менше. Середнє альбедо води змінюється від кількох відсотків у тропічних...
-
Розріз Нововолинськ?опорний розташований в кар'єрі цегельного заводу[10]. Геоморфологічно це привододільна рівнина Волинської височини. Таблиця 3.4...
-
Розріз Перемисловичі - опорний, розташований у кар'єрі цегельного заводу села Перемисловичі, Сокальського району, Львівської області, поблизу м....
-
ІІ, Підстильна поверхня. Загальна інформація - Вплив підстильної поверхні на формування клімату
Підстильна поверхня. Загальна інформація Підстильна поверхня -- сукупність природних і перетворених (змінених та створених людиною) ландшафтів на земній...
-
Поняття "охорона праці" визначено статтею1Закону України "Про охорону праці". Охорона праці - це система правових, соціально-економічних,...
-
Розріз Бояничі Розріз Бояничі - опорний [15] розташований в кар'єрі цегельного заводу у межах вододільної частини південно-західної окраїни...
-
Згідно схеми геоморфологічного районування, запропонованої П. М. Цисем [31], Волинська височина є підобластю геоморфологічної області Волино-Подільської...
-
Фауну молюсків у розрізі Коршів відібрано з середньо - та верхньоплейстоценової частини. У середньоплейстоценових лесах фауну молюсків (табл.3.8.) було...
-
Волинська височина у геоструктурному відношенні розташована у межах південно-західної окраїни Східно-Європейської платформи з двома чіткими...
-
У західній частині Волинської височини розповсюджені в основному відклади силуру. Залягають на розмитій поверхні кембрію і ордовика. Силур поділений на...
-
Різноманітність типів клімату - Вплив підстильної поверхні на формування клімату
Класифікація кліматів - це поділ кліматів Землі за різними ознаками, умовами виникнення або зв'язками з іншими географічними явищами на кліматичні зони...
-
Гидравлический разрыв пласта (ГРП) является одной из наиболее эффективных технологий интенсификации работы как добывающих, так и нагнетательных скважин....
Етапи еволюції роду Stipa і формування степів